










Introduction
During the COVID-19 pandemic, many co-infections were reported worldwide (Alanio et al., 2020; Chang et al., 2020). Among all, opportunistic fungal infections such as mucormycosis and aspergillosis are caused by common fungal molds such as Aspergillus and Mucor spp., which are responsible for a significant morbidity and mortality rate of 68% (Ventoulis et al., 2020). In the past, scientists believed that fungal infections primarily occurred in individuals with severely weakened immune systems (Song et al., 2020). However, fungal infections such as aspergillosis have increasingly been reported in patients with severe respiratory infections caused by viruses, including influenza (Gangneux et al., 2020; Lansbury et al., 2020; Hoenig et al., 2022). A recent systematic review indicated that the frequency of fungal coinfections among patients with COVID-19 was 49.7%, 23.2%, 19.8%, 6.6%, and 0.5% in Asia, America, Europe, Africa, and Australia, respectively (Seyedjavadi et al., 2022). It was reported that COVID-19 likely augmented the risk of fungal infections due to its effect on the immune system and because treatments for COVID-19 (like steroids and other drugs) can weaken the body’s defenses against fungi. These fungal infections pose a serious health risk for severely immune-compromised individuals treated with the uncontrolled use of steroids, hence termed “COVID-19-associated mucormycosis/aspergillosis” (CAM/CAA) (John et al., 2021). The indications related to aspergillosis include a running nose, stiffness, coughing with blood, pain, fever, and reduced sense of smell (Schweer et al., 2014). Symptoms associated with mucormycosis include blackish discoloration around the nose, stuffy and bleeding nose, black crusts oozing out from the nose, loosening of teeth and jaw, numbness and one-sided facial pain, swelling of the eyes, blurred vision, and problems with the respiratory system (Garg et al., 2021).
Due to the clinical limitations of fungicidal agents, like high price, inevitable toxicities, and fungal drug resistance, the search for novel antifungal drugs is a pre-requisite (Cuenca-Estrella, 2014). Previous studies have proposed that bioactive molecules that act as enzymatic antifungal inhibitors can be used as therapeutic drugs to mitigate fungal infections (Chen et al., 2020). Riboflavin, a yellow pigment also known as vitamin B2, is a key micronutrient that serves as a universal precursor to coenzymes flavin adenine dinucleotide and flavin mononucleotide. It is essential for biochemical reactions in all living cells, including bacteria and fungi (Averianova et al., 2020). Many naturally occurring flavinogenic microorganisms like bacteria and fungal species are capable of synthesizing riboflavin (Averianova et al., 2020). These flavocoenzymes also play a role in the metabolism of lipids, proteins, carbohydrates, and ketone bodies, which are sources of energy for living organisms, thereby affecting the growth and viability of living cells. Unlike animals, fungi can synthesize this essential component themselves, leading to the conclusion that targeting riboflavin production is a promising novel strategy against fungal infections (Demuyser et al., 2020). Among the various enzymes involved in the riboflavin biosynthetic pathway, riboflavin synthase (RS) catalyzes the final step in the biosynthesis of riboflavin, while 3,4-dihydroxy-2-butanone 4-phosphate synthase (DBPS) and riboflavin biosynthesis protein domain-containing protein (RibD) are major proteins implicated in the initiation of the riboflavin synthesis pathway (Jain et al., 2013). Since these proteins are essential in fungi but absent in humans, they can be effective targets for therapeutic interventions (Madhavan et al., 2022). Consequently, such proteins can serve as excellent drug target sites for anti-fungal drugs. This study postulates the viewpoint that bioactive compounds like 1,8 cineole have the potential to combat fungal infections by targeting RS, RibD, and DBPSG.
1,8-Cineole (eucalyptol), one of the most common monoterpenes, is an achiral aromatic component found in many plants, including Eucalyptus leaves. 1,8-cineole is a major bioactive component of Eucalyptus essential oil (EO) in all Eucalyptus spp. (Tyagi et al., 2011). 1,8-cineole is known for its mucolytic, broncholytic, and anti-inflammatory properties, making it useful in the treatment of respiratory tract diseases. Previous studies have documented the antifungal potential of Eucalyptus leaf extracts against Candida albicans and dermatophytes (Sebei et al., 2015). However, due to the complexity of Eucalyptus EO, its antifungal mechanism of action is still not completely understood (Elaissi et al., 2011). We present the viewpoint that due to the abundance of 1,8-cineole in EO from Eucalyptus polybrachtea it may have the potential to mitigate aspergillosis and mucormycosis. Therefore, the aim of the present study was to investigate the 3D docking of 1,8-cineole against RS, RibD, and DBPS and perform wet-lab authentication. However, the potential of 1,8-cineole against aspergillosis and mucormycosis is still a matter of speculation. This study introduces new ideas for the potential identification of key antifungal drugs during COVID-19 treatment.
Material and methods
GC-FID analysis of EO
EO was extracted from fresh leaves of Eucalyptus polybrachtea, which naturally grow in the nearby areas of Lyallpur Khalsa College, Jalandhar, India. The authentication of Eucalyptus polybrachtea was performed by Dr. Upma from Botany Department, Lyallpur Khalsa College, Jalandhar. A specimen voucher with BT103 number was deposited in the Department of Biotechnology, Lyallpur Khalsa College, Jalandhar, India. Hydrodistillation method was used for extraction of EO following the method outlined by Sebei et al. (2015). A clevenger-type apparatus (Borosil, India) was employed for the extraction process. To identify the bioactive compounds in EO, a GC-FID study was carried out (GC-FID, Chemtron 2045). The specifications of the column were as follows: 2 m long, stainless steel having 10% OV-17 on 80–100% mesh Chromosorb W (HP). Nitrogen was used as the carrier gas at a flow rate of 35 ml/min. A 0.2 μl EO sample was used. The detector and injector temperatures were set at 220 and 270°C, respectively. The oven ramping conditions were initially maintained at 100°C and ramped to 2100°C at 3°C/min. Bioactive constituents in EO were identified by comparing the relative retention times (RT) of the GC-FID spectra of the EO with authentic standards (eucalyptol, citral, eugenol and geraniol, Sigma-Aldrich (St. Louis, MO, USA)) and literature data.
Ligand prepration
For fungal receptors, including RS with Pdb id1I8D, RibD with Pdb id 7DS0, and DBPS with Pdb id 1G57, 1,8-cineole was used as a ligand. To build the 3D structure of the ligand, the SMILES notation of 1,8-cineole (PubChem CID: 2758) was retrieved from the NCBI-PubChem database. The structure was built by using UCSF-chimera. To generate the 3D structure, the SMILES notation was uploaded into the built-in tool “built structure” in UCSF-chimera, and the job was executed. The resulting file was saved as “pdb” and used for further analysis in docking.
Molecular docking
The crystal structures of RS, RibD, and DBPS fungal enzymes were obtained from PDB (https://www.rcsb.org/). Before the docking analysis, the target enzymes were prepared by removing selected H2O molecules, cofactors, and co-crystallized ligands, and undergoing energy minimization. All protein target structures were prepared by means of the dock prep set up in UCSF-chimera. The process optimized bond length, charged anomalies, and corrected atomic structure. The PatchDock tool was used for docking ligands over RS, RibD, and DBPS (https://bioinfo3d.cs.tau.ac.il/PatchDock/). Both receptors and ligand molecules were uploaded as “pdb files” to the PatchDock and docking was performed with default Clustering RMSD 4.0. The resulting complex of the topmost model with the maximum docking score generated by PatchDock was saved as “pdb” file and used for further analysis. To analyze the 3D interactions in docked complexes, the Plip tool (https://plip-tool.biotec.tu-dresden.de/plip-web/plip/index) was used. The pdb file of the docked complex was uploaded to the Plip server, and the job was executed.
Drug-likeness and toxicity
ADMET (absorption, metabolism, toxicity, and excretion), drug likeness, physiochemical properties, and pharmacokinetics were studied using the SWISSADME tool (http://www.swissadme.ch/). The pdb file of the docked complex was uploaded to the SWISSADME server, the job was executed, and the final results were retrieved. ProTox-II was used for the determination of toxicity profile, oral toxicity, hepatotoxicity, immunotoxicity, mutagenicity, carcinotoxicity, cytotoxicity, and analysis (http://tox.charite.de/protox_II). The pdb file of the docked complex was uploaded to ProTox-II server, and the job was executed. The bioactivity potential of 1,8 cineole was studied by using the web-based Molinspiration tool (https://www.molinspiration.com/cgi-bin/properties). The pdb file of the docked complex was uploaded to the Molinspiration server, and the job was executed.
Active sites prediction of proteins
To identify the cavities and determine their dimensions on the 3D active sites of the fungal receptors, the CASTp web tool was employed. All structures in “pdb” format were uploaded to the server and prediction was executed with a probe radius value of 1.4 Å. The 3D structures were viewed inside the tool, and the data was noted.
In vitro antifungal activity of EO
The antifungal activity of EO was determined using the poison food technique described by Gakuubi et al. (2017). The antiaspergillosis and antimucormucosis potential of EO were tested against the fungal strains Aspergillus niger (MTCC 277), Aspergillus oryzae (MTCC 343), and Mucor sp. (MTCC 3473). All microorganisms were bought from IMTECH (Institute of Microbial Technology, Chandigarh, India). To conduct the antifungal activity assay, the EO was added to autoclaved potato dextrose agar (PDA) agar medium (HiMedia) at concentrations of 50 μl and 100 μl. The mixture was then poured into sterile Petri plates (8 cm in diameter) and allowed to solidify at ~37°C for 3 hrs. Mycelial fungal discs (8 mm) were taken from fungal plates with the help of a sterile cork borer and aseptically placed on Petri plates. The plates were then incubated for 15 days at 29°C. As a positive control, antibiotic discs containing 25 mg ketoconazole (Leford Pvt Ltd, India) were used. MGI (Mycelial growth inhibition) was premeditated by using the following equation:
where DC and DT are the diameters of the control and test colonies. The antifungal activity experiments were performed in triplicate.
Results and discussion
GC-FID profiling
The aroma profile of EO components identified by GC-FID is displayed in Figure 1 and Table 1. GC-FID analysis of EO displayed the incidence of 39 peaks, including major and minor ones. The bioactive components identified in EO were eucalyptol (1,8 cineole) (13%), α-pinene (10%), Trans-Geraniol (7.4%), Beta-myrcene (4.5%) and citral (2.9%). As reported in the literature, Eucalyptus EO is predominant in eucalyptol (1,8 cineole) (Raho et al., 2012). Due to the presence of this bioactive molecule, eucalyptus EO has tremendous applications in health- and medical-related research (Kushwaha et al., 2018). The characteristic odor of EO oil is due to its high alcohol content, mainly 1,8 cineole, and small but varying amounts of esters associated with geraniol. Due to these bioactive molecules, EO has a rose-like aroma and has immense applications in high-grade perfumery, cosmetics, flavoring and aromatherapy, fragrances, soaps, detergents, toiletry, tobacco products, and pharmaceutical industries (Lawrence et al., 2012; Bhatnagar, 2018). Therefore, 1,8 cineole was selected as a ligand for docking studies against fungal enzyme receptors.
Table 1
Chemical composition of essential oil from Eucalyptus polybrachtea
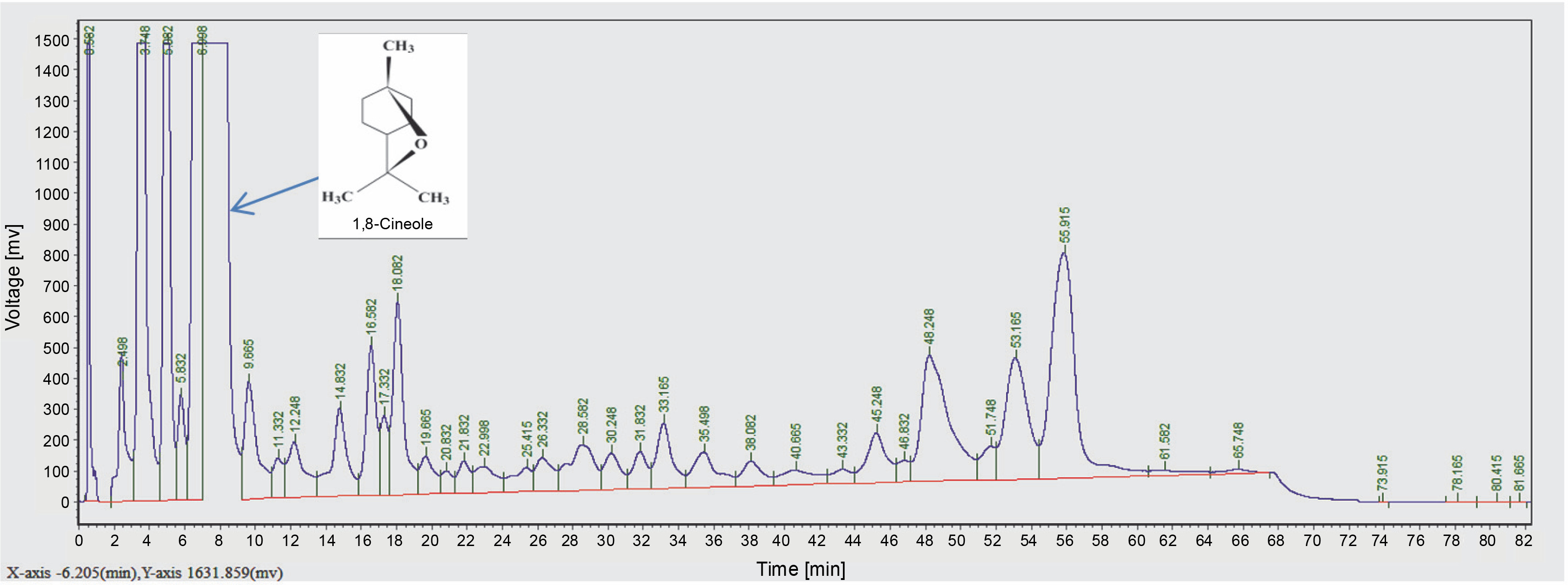
Molecular docking
Structure-based drug design (SBDD) is a widely utilized in silico technique for drug development, relying on 3-D structures (Singh et al., 2016). In silico docking has helped investigators to screen 3D conformations and possible interactions of bioactive components against receptors (Barcellos et al., 2019). The present study aimed at docking 1,8 cineole, a bioactive molecule from the EO, as a key fungal inhibitor candidate against RS, RibD, and DBPSG. From docking analysis, it was apparent that the ligand efficiently docked with RS, RibD, and DBPSG fungal enzymes. The 3D docking results depicted strong binding of fungal enzymes with 1,8 cineole (Table 2), as apparent from its docking scores. Among all receptors, 1,8 cineole depicted strong binding with RS, exhibiting a docking score of 18010. The docking score for RibD was 13132, and for DBPS it was 16790.
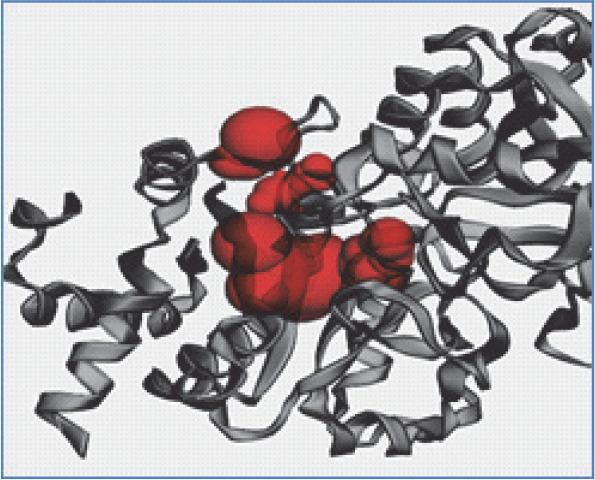
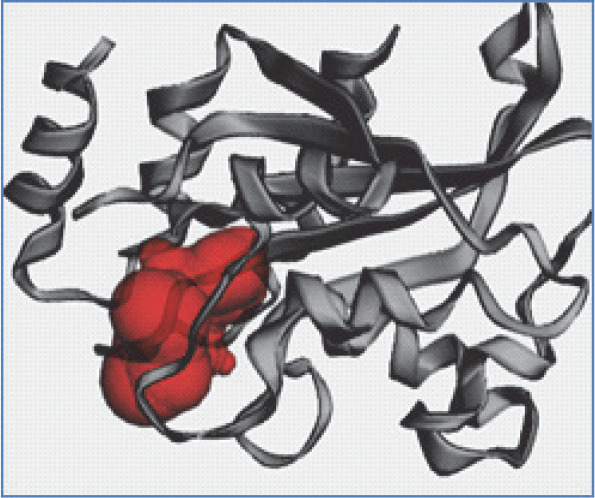
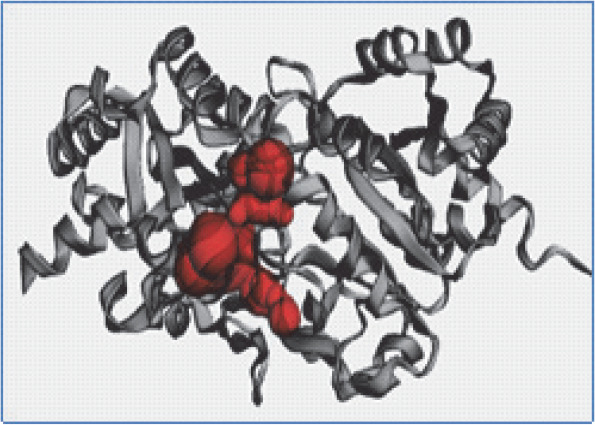
Figure 2A, Figure 2B, and Figure 2C present the 3D models depicting the docking poses, hydrophobicity view, and 2D/3D interactions of 1,8 cineole with RS, RibD, and DBPSG fungal enzymes, respectively. The hydrophobicity view reveals that ligands are firmly bound within the binding pocket of receptors. In the case of RS, 1,8 cineole docks between two monomers in the homotrimer structure of RS. With RibD, 1,8 cineole docks with the C-terminal domain of the RibD protein. These findings are in agreement with previous studies reporting docking interactions of EO-based bioactives from plants Trachyaspermum ammi, Thymus vulgaris, and Boswellia carteri with fungal enzymes such as Raslike GTP-binding protein (RHO1), Mitogen-activated protein kinase (FMK1), Histone H3-K4 methyltransferase (SET7) from Fusarium oxysporum (Biswal et al., 2019; Omar et al., 2021). The authors reported that bioactives from Thymus vulgaris EO, such as carvacrol, α-thujene, and thymol compounds, bound to the active sites of pathogenic proteins with binding affinities ranging from −4.1 to −6.9 kcal/mol. (Omar et al., 2021) Similarly, the nonacosane and tetratriacontane ligands from Boswellia carteri EO bound to pathogenic proteins with a binding energy that varied from −5.2 to −5.9 kcal/mol (Omar et al., 2021). In Trachyaspermum ammi EO, major bio-active compound ligustilide had the lowest free binding energy of −5.75 kcal/mol, followed by cedrane with −5.20 kcal/mol (Biswal et al., 2019). These studies further emphasize the potential of EO as an effective source of antifungicidal compounds.
Table 2
Molecular docking of fungal receptors with 1,8 cineole
Fig. 2A
3D model (A), hydrophobic view depicting cavity (B) and 3-D interaction (C) of RS with 1,8 cineole
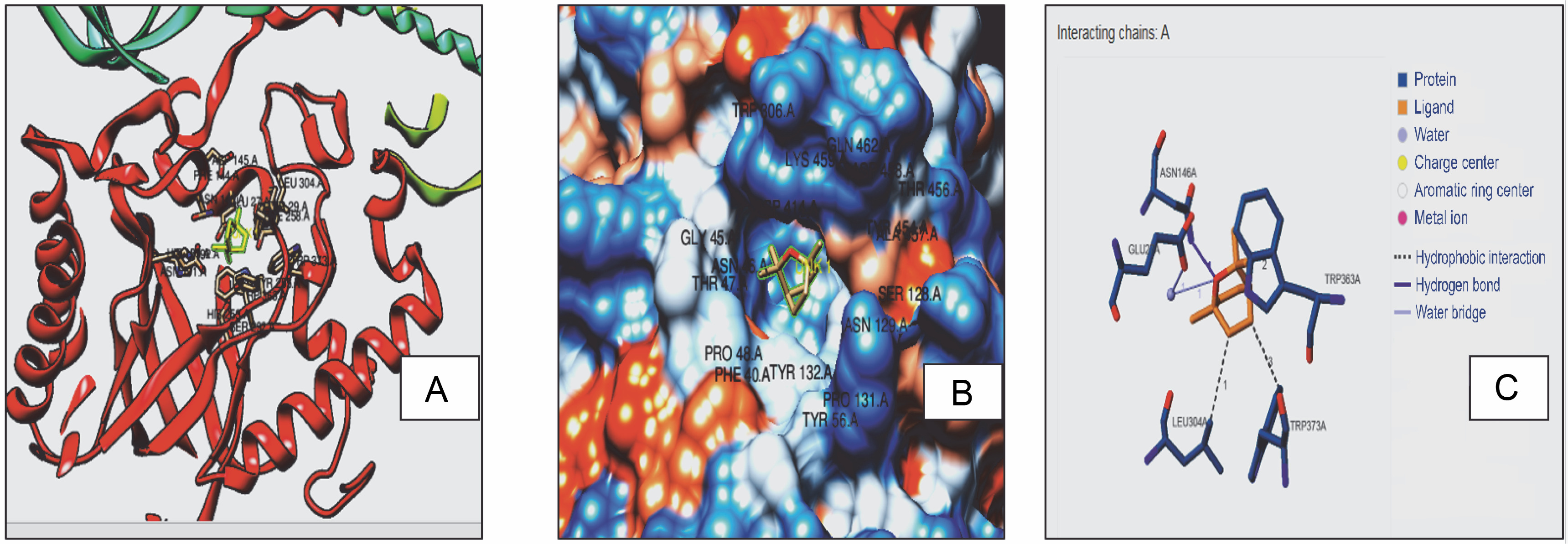
Fig. 2B
3D model (A), hydrophobic view depicting cavity (B) and 3-D interaction (C) of RiBD with 1,8 cineole
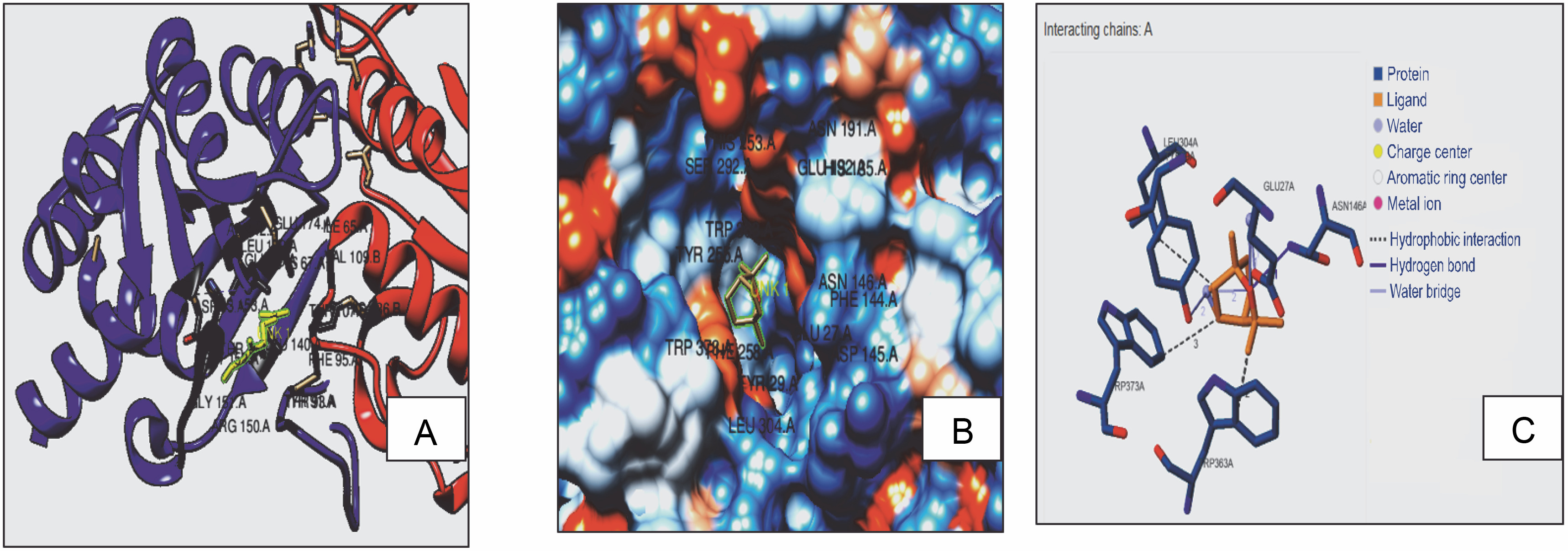
Fig. 2C
3D model (A), hydrophobic view depicting cavity (B) and 3-D interaction (C) of DBPS with 1,8 cineole
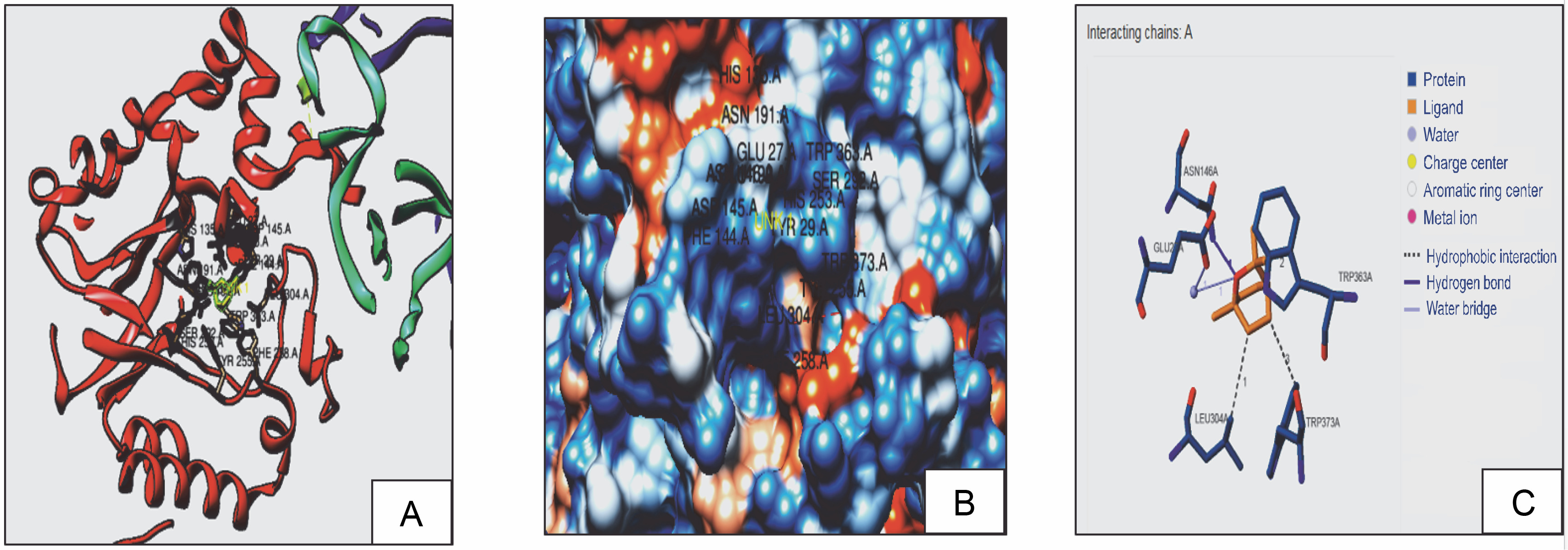
With receptors, ligands can form H-bonds or hydrophobic bonds, which designate the affinity of the ligand with the receptor (Lima et al., 2019). Hence, docking interactions of 1,8 cineole with RS, RibD, and DBPS were further evaluated. It was observed that 1,8 cineole forms both H-bond and hydrophobic interactions with RS, RibD, and DBPS. With all fungal receptors, hydrophobic interactions were detected via LEU 304, TRP 363, and TRP 373 (Fig. 2). Additionally, 1,8 cineole formed a hydrogen bond with ASN146 in all receptors CASTp active sites prediction quantified interacting residues in the active site cavities of RS, RibD, and DBPS receptors (Table 3). In RS enzymes, a main pocket was documented with a volume of 214 Å2 and an area of 376 Å2. For RibD, a volume of 158 Å2 and an area of 237 Å2, and DBPS a volume of 214 Å2 and an area of 376 Å2 were noticed. Based on these observations, it can be concluded that 1,8 cineole exhibits good affinity to all fungal enzymes. It is hypothesized that upon binding with the ligand, RS, RibD, and DBPS undergo conformational changes and become closed. All these events inhibit fungal viability, mitigating the infectivity of the fungus in the host cell. Similar in silico results highlighting hydrogen and hydrophobic interactions of bioactive components from Trachyaspermum ammi essential oil as antifungal agents have been reported. (Biswal et al., 2019). The authors described fifteen active compounds and their interactions with candidapepsin-1. Among all the compounds, ligustilide exhibited the lowest free binding energy of −5.75 kcal/mol against the candidapepsin-1, with three hydrogen bond interactions at Ile 223, Tyr 225, and Thr 222 at the active site of the enzyme followed by cedrane with −5.20 kcal/mol.
Table 3
Active site analysis of protein target structures
PASS analysis
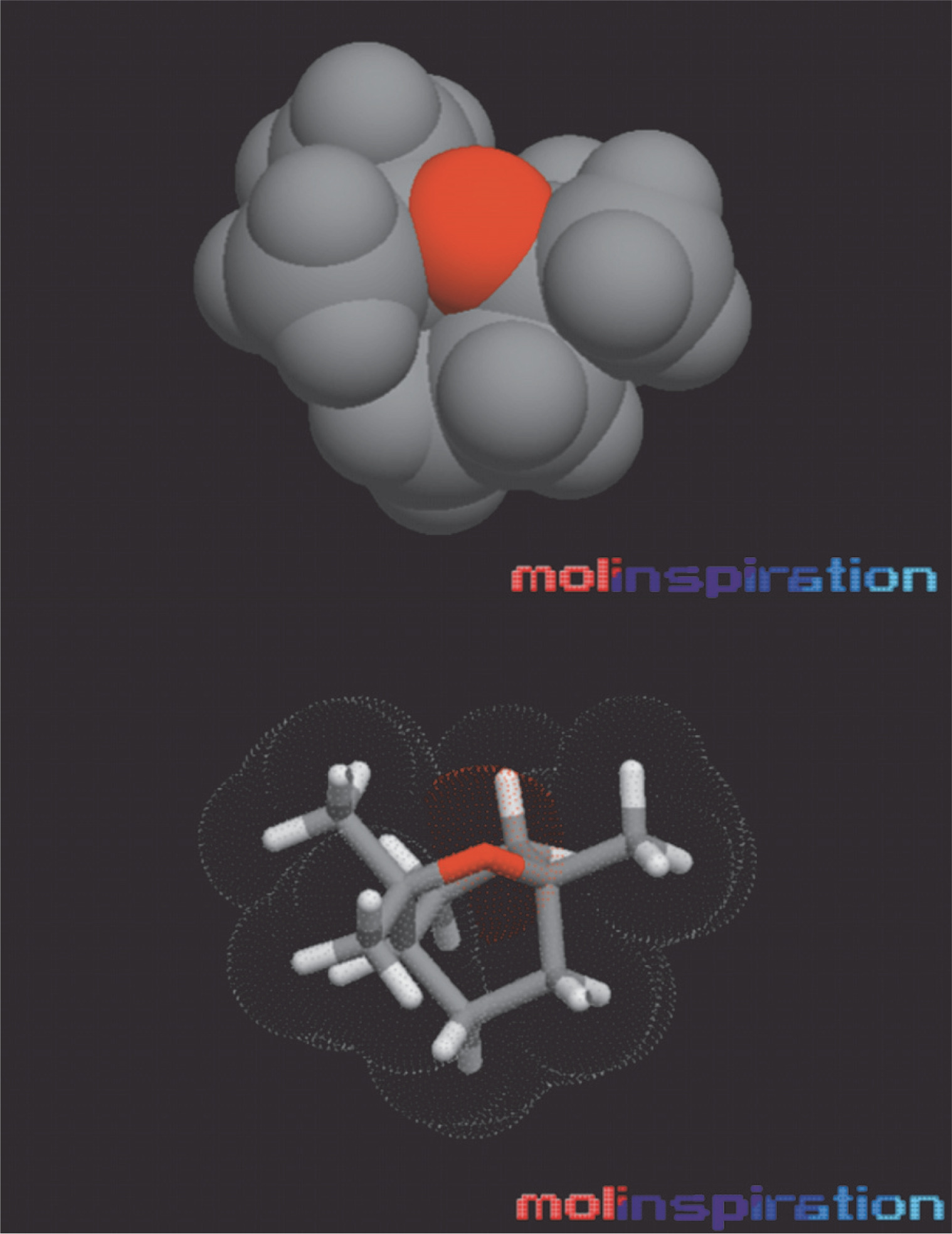
The bioactive compound 1,8 cineole underwent PASS (Prediction of Activity Spectra for Substances) and ADMET (Absorption, Distribution, Metabolism, Excretion, and Toxicity) properties analysis, which are crucial for evaluating the therapeutic potential of a drug in living organisms (Wu et al., 2020). The drug-likeness was calculated by following the Lipinski rule of five (RO5), which states that a drug must have a log P value of ≤ 5, H-bond acceptors ≤ 10, H-bond donors ≤ 5, and no more than one violation. It was observed that 1,8 cineole obeyed RO5. With 1,8 cineole, the number of H-bond acceptors was 1, and the H-bond donor was 0 (Table 4). The number of violations following RO5 was also 0. The Log P o/w value (v) for 1,8 cineole was determined to be 2.67. In pharmaco-analysis, Log P o/w is a potential factor to measure lipophilicity of any drug and its movement in the body after absorption (Abraham, 2003). Furthermore, it was observed that 1,8 cineole was a low molecular weight ligand (LMW) with a value of 154.25 g/mol. It is reported that ligands with low LMW possess a high tendency to transport across the cellular membranes and diffuse effortlessly compared to high MW compounds (Srimai et al., 2013). Molecular lipophilicity potential (MLP) depicting the surface view of eucalyptol is shown in Figure 3. The topological polar surface area (TPSA) value was 9.23 Å2, indicating that 1,8 cineole possesses good oral bioavailability (Biswal et al., 2019). TPSA is an important factor influencing drug transport properties like efficient permeability and absorption (Wu et al., 2020). To exert toxic effects, the drug has to be absorbed thoroughly into the human body. As shown in Table 4, gastrointestinal tract (GI) absorption of 1,8 cineole was high (Table 4). Additionally, it was found that 1,8 cineole was a non-substrate for P-glycoprotein (P-gp) efflux transporters. In the human body, P-gp pumps drugs back into the lumen, lessening their absorption (Konig and Muller, 2013). Moreover, as shown in Table 4, 1,8 cineole was identified as a nonsubstrate for the CYP450 series of enzymes. In the human body, CYP450 enzymes are involved in liver detoxification (Srimai et al., 2013). These results indicate that 1,8 cineole can effectively target receptors and warrant further evaluation for its biological activity score.
Table 4
Physicochemical properties and pharmacokinetics properties of 1,8 cineole
Fig. 3
Molecular lipophilicity potential (MLP)/polar surface area (PSA) views of 1,8 cineole; hydrophobic areas: encoded by violet; hydrophilic areas: red
Biological activity (BA) is a critical factor that determines the potential of a drug to bind to specific drugs or biological targets (Khan et al., 2017). In living systems, these biological targets usually include ion channels or biological receptors (Khan et al., 2017). According to the BA rule, a drug is considered active if its BA score is greater than 0, silent if it is less than −5.0, and sufficiently active if it falls between −5.0 and 0 (Khan et al., 2017). As shown in Table 5, BA scores of 1,8 cineole for various targets are as follows: −1.60 for kinase inhibitor, −1.07 for nuclear receptor ligand, −0.93 for GPCR ligand −0.90 for protease inhibitor, −0.1 for enzyme inhibitor, and 0.01 for ion channel modulator. It was observed that 1,8 cineole exhibited activity as an enzyme inhibitor and ion channel modulator. As for kinase inhibitors, nuclear receptor ligands, GPCR ligands, and protease inhibitors, the ligand demonstrated sufficient activity. These findings indicate that the studied bio-active compound possesses the necessary properties for it to potentially serve as a key drug (Khan et al., 2017). The bioactivity score provides valuable insights into the binding interactions of bioactive compounds, facilitating the development of new drugs with improved binding selectivity profiles and reduced undesired effects (Khan et al., 2017).
Table 5
Bioactivity score of 1,8 cineole with different biological targets
| Bioactivity | Score |
|---|---|
| Kinase inhibitor | −1.60 |
| Nuclear receptor ligand | −1.07 |
| GPCR ligand | −0.93 |
| Protease inhibitor | −0.90 |
| Enzyme inhibitor | −0.1 |
| Ion channel modulator | 0.01 |
The toxicity profile of 1,8 cineole was calculated using the ProTox tool. To be used as an effective therapeutic drug in pharmaceutical companies, in silico toxicity analysis is a prerequisite (Banerjee et al., 2018). Raies et al. (2016) highlighted the importance of analyzing the toxicity profile of drugs to assess potential harm to humans, the environment, or animals. The toxicity profile of the ligand was assessed, and it revealed that 1,8 cineole was nontoxic, nonmutagenic, and non-carcinogenic (Table 6). Furthermore, the bioactive compound 1,8 cineole showed inactivity toward the biological pathways-based targets such as androgen receptorligand-binding domain (AR-LBD), aryl hydrocarbon receptor (AhR), androgen receptor, weak response element heat shock factor (HSE), nuclear factor (derived from erythroid 2)-antioxidant/antioxidant response factor (nrf2/ARE), and mitochondrial membrane potential (MMP) (Table 6). Lea et al. (2017) reported that the mutagenic nature of biologically active substances is harmful to cells and is the main driver behind several diseases, such as cancer. Huang et al. (2016) reported that all these targets are principal components of biological systems inside the human body. Furthermore, the prediction results revealed that 1,8 cineole belongs to toxicity class 5, indicating the safe nature of the bioactive compound (Table 6). LD50 is the toxic unit that measures the lethal toxicity dose of the drug; the predicted LD50 value was 2100 mg/kg for 1,8 cineole. On the toxic scale, the −log10(LD50) value of 1,8 cineole was 3.3, indicating its nontoxic nature. On the poison scale, which covers a range of 13 orders of magnitude, lower −log10(LD50) values indicate the nontoxic nature of drugs (Strey, 2019).
Table 6
Toxicity profile of 1,8 cineole

In vitro antifungal activity of EO
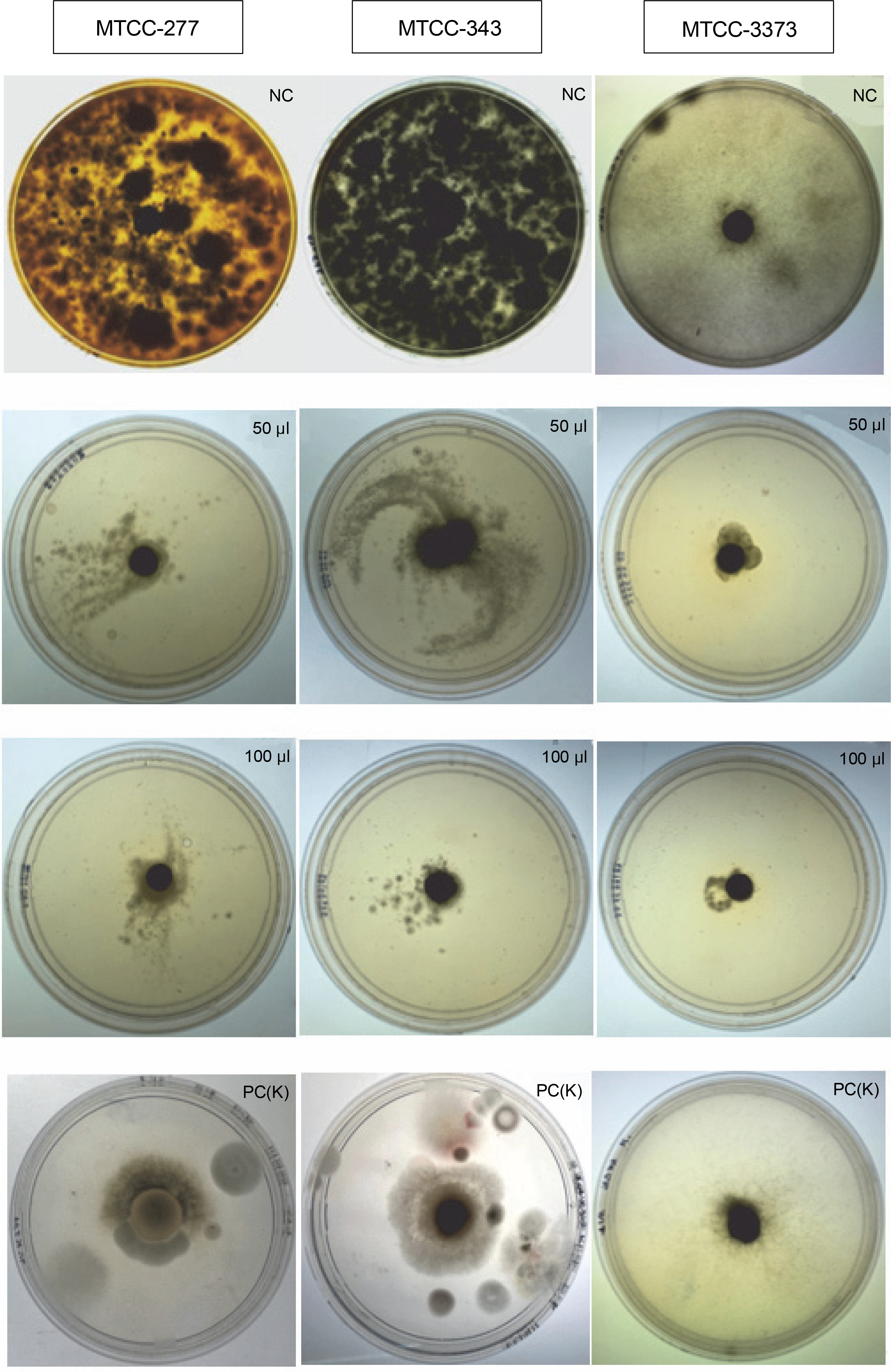
Wet lab authentication further corroborated the in silico findings by using three fungal strains, namely A. niger (MTCC 277), A. oryzae (MTCC 343), Mucor sp. (MTCC 3473). When using EO, at a concentration of 50 μl, 67% mycelial growth inhibition was observed in MTCC 3473 fungal strain, whereas 11% and 20% mycelial growth inhibition was observed for MTCC 277 and MTCC 343, respectively (Fig. 4, Table 7). At a concentration of 100 μl, the EO displayed 87% growth inhibition against MTCC 3473, 55% against MTCC 277, and 50% against MTCC 343. The antifungal potential of EO may be attributed to the presence of the major component, 1,8 cineole, or a combination of major and minor components. It is possible that the EO easily penetrates the fungal cell wall, leading to membrane leakage of electrolytes or lipid peroxidation of the membrane, which eventually hinders fungal hyphal growth and germination of conidia of fungal strains (Bakkali et al., 2008). It was also noted that certain antifungal agents inhibit fungal growth by interfering with ergosterol biosynthesis by binding to ergosterol in membranes. This damages the integrity and function of membrane-bound proteins and disturbs osmosis, fungal growth, and proliferation (Bendaha et al., 2011). Notably, this study reported 50–87% fungal inhibition by using 100 μl of EO. Fungicidal effects at a concentration of 500 μl of EO have also been pronounced for other pathogenic fungi, such as Magnaporthe grisea (81%), Botrytis cinerea (75%), Bipolaris maydis (62%), Rhizoctonia solani (46%), Colletotrichum gloeosporioides (45%), Alternuria longipes (31%), Alternaria solani (26%), and Fusarium moniliforme (25%) (Zhou et al., 2016). Similar findings have been reported by Hua et al. (2014) who studied litsea, anise, peppermint, camphor, and eucalyptus EOs and showed dose-dependent antifungal activity against Aspergillus ochraceus. The authors observed 50–70% inhibition of fungal growth rate at a concentration of 250–500 μl of oils. Passone et al. (2012) reported the effect of boldo EO against Aspergillus nigri growth and observed that boldo EO showed the greatest effect on fungal isolates with significant growth reductions at 1000 and 2000 μl, as well as total growth inhibition at 3000 μl. Matricaria chamomilla L. oil was found to be a potent inhibitor of A. niger growth in vitro, with growth inhibition of ~7.5% at 15.62 μl and reaching a maximum of ~92.5% at 1000 μl (Tolouee et al., 2010).
Table 7
Mycelial growth inhibition by EO against various fungal strains
Conclusion
The aroma profile of the EO showed that 1,8 cineole was the major component, accompanied by some other minor components. The molecular docking study revealed that 1,8 cineole from EO was an effective ligand, which docked with fungal cell wall enzymes like RS, RibD, and DBPSG. Wet-lab results showed substantial inhibition of the growth of the fungal strains that cause aspergilosis and mucormycosis diseases. Therefore, the combined findings from the in silico and wet-lab experiments established that eucalyptus plant EO might be a promising antifungal therapeutic drug against aspergillosis and mucormycosis.