










Introduction
Since antiquity, long before the advent of allopathic drugs, herbs and medicinal plants have been germane and significant in the management and treatment of many human ailments and pathological conditions (Gourhan, 1975; Falodun, 2010). Over 55,000 species of plants have been reported to be used singly or in combination in phytomedicine (Pan et al., 2014). Even with the proliferation and discovery of many conventional and synthetic drugs, plants are still recognized as important in healthcare programs and systems of many countries. Many pharmaceutical and research institutes are embracing and investing in herbs and natural products as a reliable source for drug development and discovery (WHO, 1998; Seidl, 2002; Corson and Crews, 2007; Schmidt et al. 2007; Li and Zhang, 2008).
Thaumatococcus daniellii (Benth) is a perennial, monocotyledonous, self-propagatory plant that belongs to the Marantacea family (Table 1) (Jennings et al., 2001). It is an inherently wild, shade-tolerant plant originating from sub-Saharan Africa from Sierra Leone to Democratic Republic of Congo (DRC). It is also reported to exist in Australia, Singapore, Angola, the Central African Republic, Princes’ Island, Uganda, and Indonesia (Yeboah et al., 2003). An increased demand and use of the plant for its household, presumed medicinal and industrial applications, and economic value has led to an upward spiral in its cultivation. There is no paucity of scientific data on the nutritional and pharmacological value of oils, hexane fraction, and extracts obtained from T. daniellii. Most of these data can be found in scholarly articles published in highly prestigious academic journals (Elemo et al., 1999; Adeogun et al., 2016; Shalom et al., 2018).
Table 1
Taxonomical classification of Thaumatococcus daniellii
Botanical description and taxonomy of T. daniellii
T. daniellii, also called as sweet prayer plant and African serendipity berry, is commonly referred to by its African vernacular names, including Katemfe in Sierra Leone, Nzilizili in Democratic Republic of the Congo (Termote, 2011), and Adundunmitan, Ewe eran, and Ewe moi-moi in South Western Nigeria. It is also known as Iwee and Mfang Aya among the Igala and Ibiobio tribes of North Central and South Southern Nigeria, respectively (Essien et al., 2016; Ukwubile et al., 2017). With only one species in the entire West Africa, T. daniellii is a large rhizomatous, flowering herb and grows up to approximately 40 cm in height (Hutchinson and Daziel, 1963). The fruits are pyramidal in shape and have dark red-brown hue when truly ripe (Fig. 1). At maturity, 3–4 black, hardened, and strong seeds (6–30 g) wrapped in a gummy thin, yellow basal aril are contained in the fruit (Tomlison, 1961; Most et al., 1978; Lim, 2012). Depending on the number of years lived and the habitat, T. daniellii stalks are slender and long and can grow up to 2–3 m, each carrying a lone sturdy, egg-shaped leaf (60 cm long and 40 cm wide, Fig. 2) and violet colored flowers (Makinde and Taiwo, 2004). The aril contains a nontoxic, nondiabetic, low-calorie, extremely sweet, hydrophilic protein known as thaumatin (22 000 Da) (Watson and Dallwitz, 2000). Its structure contains a single chain of 207 amino acid residues and eight disulfide bonds with an isoelectric point of 11.5–12.5 (Gloria, 2003). Antecedently, a monomeric cysteine protease (thaumatopain) of molecular weight (MW) of 30 000 Da and optimum pH 8.0 was purified from the arils (Cusack et al., 1991; Stephen et al., 1994). Furthermore, protease isolated and purified from the pulp of T. daniellii was found to have an optimum temperature of 35EC and pH 7 (Raimi et al., 2011).


Folkloric and ethnomedicinal uses of T. daniellii
Different parts of T. daniellii are used in traditional and folkloric management of some diseases and ailments. The roots and fruits are used in Democratic Republic of the Congo to treat women in labor with dystocia and childbirth complications. The seeds and leaf saps have been documented as an emetic agent in treating lung diseases and as an antidote against bee stings, snake bites, and venom, while the fruits and pulps are used in child weaning and as a laxative (Termote, 2012; Adeyemi et al., 2014; Shalom et al., 2014). Leaf and root saps are used as tranquilizing agents for treating mental and psychiatric disorders (Shalom et al., 2014). Different ethnobotanical surveys have reported the folkloric usefulness of T. daniellii in the traditional management of high blood sugar and diabetic-related disorders (Coolborn and Adegbemisipo, 2008; Olorunnisola et al., 2016). In Gabon, T. danielli is planted and cultured for its presumed fetish and magical potential (Mansfeld, 1986).
The leaves are widely used in wrapping cooked rice and moi moi during cooking in Ghana and Nigeria. The plant has also gained wide acceptance in some parts of North and South America because of its exotic and flavor enhancing property (Thorn, 2004). The petioles are used in weaving mats and baskets, whereas bakeries, local wine industries, and confectionaries use thaumatin as a flavor and taste modifier (Onwuema et al., 1979; Franke, 1985; Abbiw, 1990; Zemanek and Wasserman, 1995; Wiersema and Leon, 1999; Elemo et al., 2011). Because of worldwide popularity and international and industrial approval of thaumatin as a natural sweetener, export of the plant from West African cities and villages has led to immense economic benefits of up to US $17 000 in foreign exchange yearly (Yeboah et al., 2003; Waliszewski et al., 2005; Arowosoge and Popoola, 2006; Chiech, 2008; Boadi et al., 2014).
Phytochemistry of T. daniellii
Plants are abundant sources of phytochemicals that are needed for survival and protection against predators (Oz and Kafkas, 2017; Choudhari et al., 2020). Results from animal and cell culture studies and consumption of these phytonutrients by humans have demonstrated that antioxidants afford considerable protection against free radicals and attenuate the progression of degenerative diseases such as cancers (Chu et al., 2002; Corson and Crews, 2007; Chahar et al., 2011). Different classes of bioactive principles and secondary metabolites of plant origin have been implicated for their potential to induce a cascade of biological and pharmacological activities (Suffredini et al., 2004; Boots et al., 2008). For instance, insecticidal activities of plants such as Cymbopogon citratus, Citrullus colocynthis, Curcuma longa, Allium sativum, Cannabis indica, and Artemisia argyi have been attributed to the presence of compounds such as caryophylleneonides, α-bisabolol, dronabinol, methyl allyl disulfide, diallyl trisulfide, erucylamide, and octasiloxane hexamethyl (Ali et al., 2004; Phasomkusolsil and Soonwera, 2011; Hikal, 2017; Ahmed et al., 2020). Moreover, the importance of natural flavoring compounds such as saffron from Crocus sativus, carvone, α-lonone, eucalyptol, and menthol has been previously reported (Maggi et al., 2009; Lorenzo and Eugeniol, 2011). Alpinia gagnepainii, Phrynium imbricatum, and Hibiscus sabdariffa are among other plants that are used traditionally as food colorants and preservatives (Gyamfi and Aniya, 2002; Salavkar et al., 2011; Quang et al., 2018; Zhou et al., 2018a; Fadahunsi and Adeyina, 2019). Extensive studies have been conducted to determine the qualitative and quantitative phytochemical constituents of T. daniellii, with many studies confirming the presence of some compounds with biological importance (Table 2). Previous studies by Elemo et al. (1999), Elemo et al. (2001), Oforibika et al. (2017), and Oyeleke et al. (2017) reported that the pericarp and seeds contain high content of phytic acid, tannins, saponins, and polyphenol. Tandemly, quantitative phytochemical analyses of extracts of fruit (pericarp and seeds), leaf, and root revealed the presence of alkaloids, tannins, saponins, flavonoids, phlobatannins, anthraquinones, phytate, phenol, steroids, and terpenoids in high quantities (Adeyemi et al., 2014; Ndukwe et al., 2014; Shalom et al., 2014; Adeogun et al., 2016; Oke et al., 2016; Oboh et al., 2016; Hamid et al., 2017; Ogoloma et al., 2017). By using gas chromatography-flame ionization detector (GC-FID), gas chromatography-mass spectrophotometry (GC-MS), and high-performance liquid chromatography (HPLC), finger-printing of the essential oils and crude extract of the rhizome and leaf has shown arrays of secondary metabolites and bioactive principles (Shalom et al., 2014; Adeogun et al., 2016; Oboh et al. 2016; Ogoloma et al. 2017). Notable among these compounds are 4-hydroxy carbonyl-2-oxoethyl triphenylphosphonium bromide, 1-methyl-3-isoprophylbenzene, hendecane, 1,3,5-trime-thylbenzene, 3,3-dimethylbutyrophenone, n-nonane, isodecane, 4-methyldecane, lunamarine, ribalinidine, kaempferol, rutin, oxalate, catechin, anthocyanins, epicatechin, and sparteine (Table 3) (Shalom et al., 2014; Adeogun et al., 2016; Oboh et al., 2016; Ogoloma et al., 2017). In another study, Abiodun et al. (2014) reported that GC-MS chromatogram revealed cyclopropane, nonyl-5-tetradecene, and 7-tetradecene to constitute the highest percentage (79.60%) of the 24 compounds of essential oil of the fruit tissue and seeds of T. daniellii. Conversely, on the basis of the results of analysis with thin layer chromatography, the presence of additional compounds (cardenolides and steroidal nucleus) in the leaf crude extracts was reported (Ojekale et al., 2013). Hydrodistillation of the leaves of T. daniellii and analysis of the oil show the presence of 21 volatile constituents (Ojekale et al., 2013). A wide range of compounds such as polysiloxanes, terpenoids, aromatics, long-chain hydrocarbons, polysteroids, alcohols, aldehydes, ketones, fatty acids, and their esters were discovered using GC and GC-MS analysis, with octadecamethyl cyclononasiloxane (48.3%), phytol (12.9%), and 6,10,14-trimethyl-2-pentadecanone (9.3%) being the dominant compounds (Ojekale et al., 2007). In congruence, GC-MS analysis also recorded the bioavailability of 22 compounds in the essential oil of the leaf, among which are oleic acid and geranylgeraniol (Omisope et al., 2016). Compounds such as squalene, tetracontane, n-hexadecanoic acid, phytol, L-ascorbic acid, and octadecanoic acid (Table 3) were discovered through GC-MS examination as constituents of the hexane, ethyl acetate, and methanol extracts of the leaf (Hamid et al., 2017).
Table 2
Biological and pharmacological activities of some identified compounds from T. daniellii
| Compounds | Biological activity | References |
|---|---|---|
| Anthocyanins | anti-radical, anti-hyperglycemic, anticancer | Huang et al. (2014); Lee et al. (2015); Samadder et al. (2017); Thibado et al. (2018) |
| Anthraquinones | anticancer, anti-acne, antifungal, antibacterial, anti-inflammatory, anticoagulant, neuro therapeutic | Huang et al.(2007); Gan et al. (2008); Wuthi-udomlert et al. (2010); Khan et al. (2011); Seo et al. (2012); Jackson et al.(2013); Shrestha et al.(2014); Shrestha et al. (2015) |
| Catechins | anti-cancer, anti-inflammatory, antioxidant | Murakami et al. (2002); Fang et al. (2003); Hussain et al. (2005); Fang et al. (2007) |
| Geranylgeraniol | testosterone and peroxisome proliferator-activated receptor γ (PPARγ) enhancement | Ho et al. (2016); Mastubara et al. (2018) |
| Hexadecanoic acid | cytotoxic | Ravi and Krishnan (2017) |
| Kaempferol | cytotoxic, antioxidant, anti-inflammatory | Leung et al. (2007); Hamalaina et al. (2007); Vellosa et al. (2011); Boadi et al. (2016) |
| Phytol | antinociceptive, antioxidant | Santos et al. (2013) |
| Rutin | cardio-protective, neuro-protective, antidiabetic, nephroprotective | Stanley and Kamalakkannan (2006); Yang et al. (2008); Annapuma et al. (2009); Gulpinar et al. (2012); Hao et al. (2012) |
| Saponins | hypocholesterolemic, immunodulatory; anti-inflammatory, cytotoxic | Matsuura (2001); Sun et al. (2014); Lin et al. (2016) |
| Spartein | hypoglycemic; anti-convulsant | Kozlowska et al. (2007); Vargas and Ceja (2016) |
| Squalene | anti-hyperlipidemia, antioxidant, anti-atherosclerosis | Motawi et al. (2010); Gabas-Rivera et al. (2015); Kumar et al. (2016) |
| Terpenoids | anticancer, antibacterial | Dorman and Deans (2000); Huang et al. (2012) |





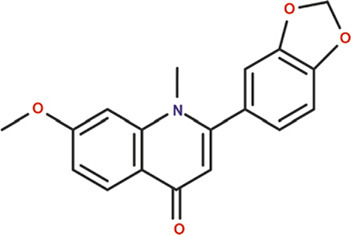


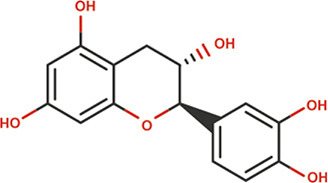


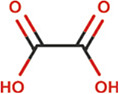
Proximate and nutritional composition of morphological organs of T. daniellii
There is overwhelming scientific information on the nutritional composition of various morphological organs of T. daniellii, which can be correlated to its nutritional and medicinal benefits. A pioneer study by Adesina and Higginbotham (1977) confirmed that arabinose, glucoronic acid, xylose, and 4-O-methyl glucoronic acid exist in the polysaccharide gel extracted from the fruit of T. daniellii. Furthermore, extensive elemental analyses have shown that many macro- and microelements are present in the different organs of the plant (Shalom et al., 2014; Sotande and Oluwadare, 2014). It was revealed that the stalk of T. daniellii contains phosphorus and nitrogen (Sotande and Oluwadare, 2014). The contents of potassium, iron, manganese, and copper were revealed using flame atomic absorption spectrometer (AAS) (Shalom et al., 2014; Sotande and Oluwadare, 2014). Potassium and iron were considered as the main metals contributing to the physiological development of the plants (Shalom et al., 2014; Sotande and Oluwadare, 2014). It has been shown that the arils, fruits, and leaves are good sources of sodium, calcium, magnesium, and phosphorus (Shalom et al., 2014, Ojo et al., 2017, Osuocha et al., 2018). Phosphorus (0.37 g) and calcium (0.34 g) were reported to be the most abundant elemental constituents per 100 g of the extract. On the other hand, the concentrations of iron and zinc were low in the leaves and fruits, respectively. Vitamins (A, B1, B3, B5, B6, B12, and C) have been reported to be present in the leaf and seed extract of the plant, with vitamin B12 as the most abundant one (Abiodun et al., 2014; Shalom et al., 2014; Oforobika et al., 2017; Osuocha et al., 2018). Proximate composition of 100 g per sample of aqueous extract of the leaf, fruit, and seed depicted a high content of carbohydrate (29.08%) in the seed and fruit, while crude fiber was more abundant in the leaf (36.61%). Fat (3.85%), protein (19.01%), and ash (10.60%) were also present, while the highest moisture (83.90%) content was detected in the seed extract (Ndukwe et al., 2014; Oforobika et al., 2017). Interestingly, proximate composition recorded for T. daniellii morphological organs were similar to that of some selected edible leafy vegetables (Table 4). Nutritional analysis of the fruit revealed that it contains calcium (6.68%), magnesium (0.60%), ash (5.80%), fiber (0.41%), protein (19.01%), nitrogen (3.04%), and hydrogen cyanide (0.81/kg) (Ndukwe et al., 2014). Defatted seed flour of T. daniellii was recorded to contain crude fiber (36.92%), carbohydrate (40.07%), and ash (8.17%), while oleic (42%), palmitic (24%), and linoleic (18%) acids were the main fatty acid constituents of the seed oil (Abiodun et al., 2014). To increase abundance, carbohydrate (44%), protein (33%), moisture (12%), fiber (5%), ash (4.8%), and fat (0.61%) were reported as the proximate composition of the aril of T. daniellii (Sotannde and Oluwadare, 2014). Fiber property examination of the stalk revealed that holocellulose (66.98%) was the most dominant component, while α-cellulose (38.51%), crude fiber (32.61%), lignin (13.29%), and hemicellulose (28.20%) were present in appreciable amounts. However, ash (2.79%) and silica (SiO2) (0.85%) were found to be present in very low concentrations (Sotannde and Oluwadare, 2014).
Table 4
Comparative proximate composition of some edible leafy vegetables with T. daniellii
| Plant | Ash [%] | Carbohydrate [%] | Fat [%] | Moisture [%] | Crude fiber [%] | Protein [%] | References |
|---|---|---|---|---|---|---|---|
| T. daniellii (Fruit) | 5.80 | 70.00 | 3.85 | 83.90 | 0.42 | 19.01 | Ndukwe et al. (2004) |
| T. daniellii (Seed) | 8.17 | 40.07 | 2.32 | 10.39 | 36.92 | 2.14 | Abiodun et al. (2014) |
| T. daniellii (Aril) | 4.79 | 44.17 | 0.61 | 12.20 | 5.20 | 33.03 | Ojo et al. (2017) |
| T. daniellii (Leaf) | 10.60 | 31.25 | 0.90 | 5.16 | 36.61 | 14.88 | Oforobika et al. (2017) |
| T. daniellii (Rhizome) | 6.20 | 18.75 | 1.80 | 17.27 | 48.98 | 7.00 | Oforobika et al. (2017) |
| Solanium macrocarpon | 8.44 | 49.56 | 1.31 | 49.53 | 11.83 | 21.54 | Odoh et al. (2007) |
| Moringa olifera | 5.13 | 43.78 | 2.63 | 71.73 | 19.25 | 20.51 | Sharma et al. (2012) |
| Amaranthus hybridus | 16.43 | 25.56 | 2.31 | 53.53 | 10.93 | 18.84 | Odoh et al. (2007) |
| Amaranthus viridis | 1.85 | 7.67 | 0.47 | 87.90 | 1.93 | 2.11 | Sharma et al. (2012) |
| Vernonia amygdalina | 12.94 | 35.56 | 4.31 | 49.53 | 9.83 | 31.54 | Odoh et al. (2007) |
| Gongronema latifolium | 16.74 | 39.56 | 1.31 | 49.53 | 11.83 | 29.54 | Odoh et al. (2007) |
| Ocimum gratissimum | 27.44 | 40.56 | 2.91 | 31.53 | 15.03 | 41.54 | Odoh et al. (2007) |
| Murraya koenigii | 12.04 | 56.56 | 6.21 | 49.53 | 4.43 | 13.34 | Odoh et al. (2007) |
Elemo et al. (1999) reported that the extracted oil from pericarp and seed was golden yellow in appearance and slightly solid at room temperature. The iodine number of the seed oil (20.5 g/100g) and pericarp oil (34.1 g/100g) was lower than that of palm oil (48–56 g/100g) and olive oil (80–88 g/100g) but higher than that of palm kernel (14.23 g/100g) and butter (26.42 g/100g). Conversely, the saponification value of the pericarp and seeds oils was lower (154.4 mgKOH/g and 149.3 mgKOH/g, respectively) than that of palm oil (196–202 mgKOH/g), coconut oil (250–264 mgKOH/g), and palm kernel (245–255 mgKOH/g). Free fatty acid composition of the pericarp and seed oil (16.2% and 14.5%) was lower than that of palm oil (37.6%) and similar to that of palm kernel (14.6%) (Weiss, 1983; Elemo et al., 1999). The refractive index, specific gravity, acid, saponification, peroxide, iodine, and free fatty acid value of the seed oil were reported to be 1.4, 0.76, 44.93 mg/g, 190.08 mg/g, 11.00 mg/g, 60.34 mg/g, and 22.58 mg/g, respectively (Abiodun et al., 2014). In addition, from the results obtained from biological evaluation, it was inferred that the nutrient enhancing ability of the seed waste proteins was comparable to that of casein (Elemo et al., 2011).
T. daniellii fruit improved the physicochemical and sensory property of “Kunun-Zaki” fermented nonalcoholic sorghum-based drink and also enhanced the shelf life of stored orange juice (Adeogun et al., 2017; Ojo et al., 2017).
Pharmacological and biological activities of T. daniellii
Antioxidant activities of T. daniellii
Increased production of reactive oxygen species (ROS) leading to disruption or an imbalance in the antioxidant defense system is referred to as oxidative stress (Stamati et al., 2011; Bhattacharya, 2015). Free radicals such as superoxide radical (O2−) and hydrogen peroxide (H2O2) are byproducts of mitochondrial electron transport and other metabolic processes (Shinde et al., 2012). They are, however, generated in high amounts from exogenous factors such as air pollution, cigarette smoking, excessive alcohol consumption, and ultraviolet radiation (Lobo et al., 2010). ROS have been implicated in the pathophysiology of several diseases such as cancer, atherosclerosis, and Parkinson’s and Alzheimer’s disorders (Parihar et al., 2009; Simone et al., 2010). Edible vegetables and medicinal plants are known to contain many secondary metabolites that can quench free radicals, upregulate the expression of enzymatic antioxidants, and thereby protect the cells and important biological molecules from oxidative damage (Baodi et al., 2006; Adegbola et al., 2020).
According to Shalom et al. (2018), the daily administration of ethanol leaf extract (500–1500 mg/kg) for 14 days showed a significant (P < 0.05) increase in the concentration of liver and kidney superoxide dismutase (SOD) and reduced glutathione (GSH) in treated rats as compared to that in the control group. An in vitro antioxidant assay revealed that phenolic extract (0–5 mg/ml) of the leaves caused a significant (P < 0.05) dose-dependent Fe2+ chelation, lipid peroxidation, and malondialdehyde (MDA) production inhibitory activities. In the same study, a dose-dependent DPPH (1,1-diphenyl-2-picrylhydrazyl) and hydroxyl radical (OH*) scavenging activity with EC50 of 0.56 and 0.76 mg/ml, respectively, was reported. According to Oboh et al. (2016) and Hamid et al. (2017), n-hexane and ethyl acetate extract (1.95–1000 μg/ml) of the leaves exhibited nonsignificant DPPH radical scavenging activities; however, the methanol extract (1.95–1000 μg/ml) inhibited 50% DPPH radical scavenging activity at the concentration of 615.14 mg/ml, which was statistically comparable to that of the control (ascorbic acid). In another study, DPPH and ferric reducing antioxidant potential (FRAP) of hexane and aqueous fractions of leaves at the concentration of 100–500 mg/ml were evaluated (Anyasor et al., 2016) The aqueous extracts demonstrated a significantly higher DPPH and FRAP inhibitory activity than hexane fraction (Anyasor et al., 2016). The observed higher activity is due to the higher phytochemical content in aqueous leaf fraction reported by Oke et al. (2016). Adedosu et al. (2017) reported that the aqueous leaf extract of T. daniellii at 100 mg/kg upregulated the antioxidant status of streptozotocin-induced diabetic rats. A significant increase was observed in GSH, SOD, and catalase activities in the extract-treated animals. Simultaneously, malondialdehyde (MDA) concentration was observed to be nearly the same when compared with that in the control and standard drug (glibenclamide)-treated animals. However, MDA was significantly lower than that in streptozotocin-induced untreated diabetic animals (Adedosu et al., 2017). Thus, this indicated the capacity of the plant extract to improve antioxidant defense and attenuate oxidative stress.
The ability of the essential oil of T. daniellii leaves to scavenge DPPH radicals and nitric oxide has been documented by Ojekale et al. (2013). The essential oil significantly and dose-dependently reduced the amount of DPPH radicals. At the concentration of 50 mg/l, more than 80% of DPPH radicals were inhibited (IC50 value of 26.3 mg/l). This observed activity was relatively higher than that of butylated hydroxyanisole, butylated hydroxytoluene, ascorbic acid, and α-tocopherol with IC50 values of 36.1, 35.9, 36.8, and 41.7 mg/l, respectively. In addition, a dose-dependent nitric oxide scavenging activity (78.5%) higher than that of ascorbic acid and α-tocopherol was shown by the essential oil. It was inferred that the essential oil of T. danielli quenches free radicals and is a good source of natural antioxidants, which can prevent lipid peroxidation, oxidative stress, and metabolic disorders.
Nwonuma et al. (2016) evaluated the effect of T. daniellii leaf extract in rats exposed to a single daily dose of potassium bromate (KBrO3) (0.5 ml of 10 mg/kg) for 2 weeks. It was reported that treatment with different concentrations of the leaf extract protected against cellular degeneration of the testes and buoyed antioxidant enzyme activities. Furthermore, there was a 20% decrease in MDA concentration of the leaf extract-treated animals, and a significant increase in GSH and SOD activities was also recorded.
Shalom et al. (2017) concluded that T. daniellii seeds are a source of natural antioxidant and may be used further for their pharmacological potential. Their study showed that 14 days administration of 500, 1000, and 1500 mg/kg seed ethanol extract of T. daniellii caused an increased hepatic SOD and GSH concentration. No significant effect on renal SOD activity and GSH concentration was observed, but a significant (P < 0.05) increase in renal GSH concentration was demonstrated at 1500 mg/kg extract concentration. T. daniellii seed had no effect on the level of hepatic thiobarbituric acid reactive substances (TBARS). However, a significant increase was observed in renal TBARS level. The authors concluded that the observable increase in SOD (28.17 U/mg protein) and GSH (20.32 mM/mg protein) after the administration of T. daniellii seeds may be due to the synergistic stimulatory effects of various active components present in the seeds, while the increased renal MDA content might be related to high oxalate concentrations in the seeds (Noonan and Savage, 1999).
Hepatoprotective activity and toxicological studies on T. daniellii
The recent surge in the use of medicinal plants and products, especially in developing nations, is based on the premise that herbal medicine is readily available, cheap, and less toxic. A noteworthy finding is that the consumption of medicinal plants at high doses can induce injury to vital organs such as the liver and kidney (Chikezie et al., 2015). Biochemical parameters such as alanine aminotransferase (ALT), aspartate aminotransferase (AST), total bilirubin, and alkaline phosphatase (ALP) in the blood can be measured to determine the functional integrity of the liver (Satyanarayana and Chakrapani, 2007). Significant changes in the concentration of AST, ALT, and ALP singly or in combination may suggest physiological changes in response to toxic plant extracts or materials (Howida, 2016; Mumoli et al., 2016). Additionally, the toxicity of many compounds is commonly extrapolated from the concentration that can cause 50% mortality in the tested population (median lethal dose or LD50). Considering the aforementioned risks associated with the indiscriminate consumption of medicinal plants, various extracts and fractions of morphological organs of T. daniellii were evaluated in many toxicological studies. As reported in the study of Ogoloma et al. (2007), a 4-week intraperitoneal administration of the methanol leaf and root extract of T. daniellii at 100-500 mg/kg doses in Wistar rats revealed the LD50 value to be 330 and 250 mg/kg, respectively. Findings from this study showed that the methanol leaf extract of T. danielli in low and high doses had no significant effect on AST. Furthermore, a statistically nonsignificant elevation in ALT level was observed. There was an observable nonsignificant change in conjugate bilirubin, which was suggestive of intact liver function. However, a noticeable increase in alkaline phosphatase (ALP) activity was observed after 4 weeks of leaf extract administration. However, Emudainohwo et al. (2015) reported an LD50 value of above 6000 mg/kg for the leaf extract of T. daniellii.
Okafor et al. (2019) examined the hepatotoxic characteristic of the aqueous and methanol leaf extract (200 mg/kg). The aqueous extract caused a nonsignificant increase in the activity of ALT (5.43 IU/l), AST (16.93 IU/l), and ALP (160.70 IU/l). In contrast, the methanol extract caused a significant elevation of ALT (13.15 IU/l), AST (22.84 IU/l), and ALP (170.40 IU/l) levels, and the histological observation of the liver tissues showed inflamed hepatocytes with enlargement of the central vein, which was suggestive of derangement in the normal architecture of the liver tissue. Hence, Okafor et al. (2019) suggested that drug development protocols involving T. daniellii leaf should preferably use water as an extracting solvent.
As stated by Iheagwam et al. (2017), a significant decrease in the AST concentration of ethanol seed extract (1500 mg/kg)-fed rats was observed. Despite this high dose exposure, a nonsignificant effect on total bilirubin (17.73 μmol/l) and direct bilirubin (26.41 μmol/l) concentration was concomitantly observed. The extract showed a better effect on these indices when compared with the standard drug (vitamin C). Hepatoprotective ability of the seed extract was further corroborated from the histological analysis, which revealed an absence of nontoxic symptoms and intact liver membrane integrity. In addition, Nwonuma et al. (2016) reported an absence of testicular cellular degeneration in the leaf extract-treated animals compared to that in the KBrO3-treated group. Moreover, the results obtained from the daily administration of 500 mg/kg acetaminophen for 4 weeks indicated that the leaf methanol extract-treated animals exhibited a significantly reduced total albumin (0.80 mg/dl), bilirubin (4.87 g/dl), AST (29.93 u/l), ALT (28.90 u/l), and ALP (115.65 u/l) as compared to those in the untreated animals. Furthermore, histopathological examination showed that the oral administration of fruit pulp ethanol extract exhibited no pathological abnormality on rat liver tissue (Iheagwam et al., 2017). Ukwubile et al. (2017) therefore concluded that T. daniellii may contain metabolites that are capable of ameliorating or protecting against liver injury.
According to Olorunnisola et al. (2017), the LD50 value of ethanol leaf extract was reported to be greater than 5000 mg/kg in male Wistar rats. In the same study, the nephroprotective effect of ethanol leaf extract of T. daniellii on streptozotocin-induced diabetic rats was evaluated. Oral administration of 250 and 500 mg/kg of the extract dose-dependently ameliorated deranged electrolyte balance and compared favorably with the effect of glibenclamide (Olorunnisola et al., 2017). Lethality assessment of the hexane, aqueous acetone, and ethanol leaf extract (20, 40, 60, 80, and 100 μg/ml) against larvae of brine shrimp naupli revealed that the aqueous and ethanol extracts of the leaf were nontoxic with LD50 values of 592.95 and 281.12 μg/ml, respectively. However, acetone and hexane extracts showed significant toxicities (LC50 of 31.27 μg/ml and 22.77 μg/ml, respectively). The discrepancies in toxicity of the extracts are suggestive of the presence of cytotoxic and harmful compounds, which might be attributed to the extraction solvents (Adeogun et al., 2016).
Hypolipidemic activities of T. daniellii
Hypolipidemic activities of ethanol leaf extracts of T. daniellii (500–1500 mg/kg) were evaluated by Shalom et al. (2018). The study revealed that a 2-week administration of the extracts had a positive effect on the lipid profile of the plasma, kidney, and liver. A significantly lower cholesterol concentration was observed in the plasma of animals treated with 1500 mg/kg extract. Plasma low-density lipoprotein (LDL) level was also considerably reduced to 0.53, 0.47, and 0.48 mmol/l for all extract concentrations (500, 1000, and 1500 mg/kg, respectively) as compared to control (0.62 mmol/l). A significantly increased high-density lipoprotein (HDL) concentration was observed in the lipid analysis of the plasma (1.64 mmol/l), liver (0.18 mmol/l), and kidney (0.05 mmol/l) of 1000 mg/kg extract-treated animals and were respectively comparable to 1.12, 0.12, and 0.05 mmol/l observed for the vitamin C-treated group.
Juxtaposing the serum lipid analysis of the aqueous leaf extract-treated rats with streptozotocin-induced untreated diabetic rats, it was observed that in the 100 mg/kg extract-fed group, serum cholesterol (72.66 mg/dl) and triglycerides (265.02 mg/dl) levels were significantly reduced compared to 120.25 and 386.66 mg/dl, respectively, noted in untreated diabetic rats (Adedosu et al., 2017). In addition, serum HDL values of 30.90 and 19.00 mg/dl were recorded for extract-treated and untreated diabetic rats, respectively (Adedosu et al., 2017). Osuaocha et al. (2018) reported that ethanol and aqueous leaf extracts (200 mg/kg) exhibited no adverse effect on the lipid profile of rats. The measured values were not significantly different from those of control animals treated with distilled water. As mentioned by Omisope et al. (2016), the essential oil extract of leaf conferred adaptogenic ability and capacity to tolerate nonspecific stress and restored or maintained normal lipid homeostasis in stress-induced hyperlipidemic rats. It was concluded that the extract reduced serum LDL concentration from 28 to 15 mg/dl and concomitantly increased HDL level from 13 to 25 mg/dl.
Antimicrobial activities of T. daniellii
Recently, there has been a worldwide drive and resurgence to develop antimicrobial agents from natural sources, which would serve as an alternative to the presently available synthetic drugs. Residues of synthetic antibiotics have been reported as a source of pollutants in animal feeds and in aquatic and terrestrial ecosystems (Kim and Aga, 2007; Kraemer et al., 2019). Several cases of antimicrobial drug resistance due to prolonged and indiscriminate use of antibiotics have been reported in the literature (Manaia, 2017; Zhou et al., 2018b). Many studies have reported various levels of efficacy of medicinal plants (Fadahunsi et al., 2017; Fadahunsi and Adeyina, 2019; Badmus et al., 2019 Olasehinde et al., 2019; Adegbola et al., 2020). Different solvent extracts and fractions of T. daniellii have been evaluated against various species of microorganisms, and the obtained results indicated that the plant might be a good candidate for the development of antimicrobial agents from natural origin. By using the disc diffusion method, Adeogun et al. (2016) evaluated the antifungal properties of acetone, ethanol, hexane and aqueous extracts of T. daniellii at the concentrations of 0.1-10 g against isolated fungi (Aspergillus aculeatus, Aspergillus niger, Aspergillus flavus strain, Rhizopus stolonifer, Issatchenkia orientalis, Meyerozyma guilliermondii, Fusarium oxysporum, Paecilomyces variotii, Penicillium crustosum, Trichoderma harzianum, and Meyerozyma caribbica) from orange juice and corn jell-O (Eko ). It was observed that the ethanol extract showed the most potent activity against the tested isolates. Moreover, the ethanol and acetone extracts exhibited the lowest minimum inhibitory concentration (MIC), highest fungicidal, and zone of inhibition activities in a dose-dependent manner. In contrast, the aqueous and hexane extracts were quite ineffective. In the study of Adebayo and Kolawole (2010), the broth dilution method showed the sensitivity of spoilage fungi (Saccharomyces cerevisiae, S. chevalieri, Rhizopus, Mucor, Penicillium, Aspergillus niger, and Aspergillus flavus) isolated from Eba, an indigenous staple food in Nigeria, and white bread to aqueous and methanol leaf extract of T. daniellii. It was noted that the methanol extract of T. daniellii had greater activity against the tested organisms than the aqueous extract.
By using the agar well diffusion method, the antibacterial activity of ethanol leaf extract (1.25–20 mg/ml) of the plant was investigated against gram-positive Staphylococcus aureus ATCC 29213, Bacillus subtilis ATCC 12432, and Streptococcus pyogenes ATCC 8668 and gram-negative Shigella dysenteriae (clinical isolate), Campylobacter jejuni (clinical isolate), and Salmonella typhi ATCC23564. The antibacterial activity exhibited by the extract was considerable and comparable to that of ciprofloxacin (Ukwubile et al., 2017). Interestingly, chloroform, methanol, ethanol, and aqueous leaf extracts exhibited no activity against the selected strains of food spoilage microorganisms (Salmonella typhimurium, Shigella dysenteriae, Shigella sp., Escherichia coli, Staphylococcus aureus, Streptococcus lactis, Leuconostoc sp., Pediococcus cerevisiae, Bacillus cereus, Candida krusei, Candida albicans, Aspergillus niger, Aspergillus flavus, and Trichoderma konigii ) (Adegunloye et al., 2006; Ojekale et al., 2007).
Antidiabetic and antihyperglycemic activities of T. daniellii
Medicinal herbs have been popularly used for managing glucose-related metabolic disorders in many developing nations. This might be due to continued ancestral practices, beliefs, and dispositions; ubiquitous nature of these plants; financial constraints due to low income; and unwanted side effects of prolonged use of allopathic and conventional diabetic medications (Sofowora, 1993; Kubukeli, 1999; WHO, 2002; Parmer, 2005; Tabut, 2006; Anoka, 2012; Das and Barman, 2012; Priscilla, 2013). According to Trinter (2007), Africans have developed their own effective ways of treating diseases long before the arrival of conventional medicine. Different ethnobotanical surveys have reported the use of T. daniellii plant parts as a recipe in the formulation of traditional and folkloric antidiabetic agents (Coolborn and Adegbemisipo, 2008; Borokini et al., 2013). Some researchers have also reported different glucose lowering abilities and mechanisms of action of the plant. According to Elemo et al. (2000), three glycoproteins (P1, P2, and S) isolated and purified from T. daniellii seed and pulp crude extracts conferred considerable inhibitory activity against α-amylase. The highest inhibitory activity (57%) observed was demonstrated by glycoprotein of the seed with optimum temperature and pH of 30EC and 6.5, respectively.
Antihyperglycemic activities of methanol and ethanol leaf extracts of T. daniellii in streptozotocin-induced diabetic rats have been reported (Chikezie et al., 2015; Ogoloma et al., 2017). Adedosu et al. (2017) reported that fasting blood sugar (FBS) of extract-fed diabetic rats (145 mg/dl) was significantly lower than that of the un-treated diabetic rats (450 mg/dl) after 15 days of treatment. To clarify the likely mechanisms of action, Olorunnisola et al. (2019) evaluated the in vitro antidiabetic activity of ethanol leaf extract of T. daniellii. It was observed that the ethanol leaf extracts (250, 500, and 750 μg/ml) induced a dose-dependent α-amylase inhibition (38.41, 46.31, and 50.81%, respectively). An increased glucose uptake in yeast cells was also observed after 72 h, which was better than the standard drug metronidazole. Simultaneously, T. daniellii showed better (87.51%) ability to inhibit hemoglobin glycosylation than standard drug (82.10%) (trolox). In the same study, in vivo glucose lowering activity showed that the administration of the extract at 250 and 500 mg/kg body weight resulted in a significant (P < 0.05) dose-dependent decrease in the blood glucose as compared to the diabetic control group. In addition, Emudainohwo et al. (2015) reported that acute and sub-acute treatments with ethanol leaf extract (200 and 400 mg/kg) caused a significant reduction in fasting blood glucose value and glycosylated hemoglobin (HbA1) and an observable increase in serum amylase of diabetic animals.
Effect of T. daniellii extracts on hematological parameters
Investigation on the effect of methanol leaf and rhizome extracts on hematological parameters showed a reduction in red blood cell and white blood cell count. Animals with low white blood cells are highly susceptible to develop infection and are incapable of generating antibodies for disease resistance (Soetan et al., 2013). However, no significant effect (P < 0.05) on hemoglobin, hematocrit, and platelet concentrations within the 28-day study period was observed. It was also noted that both extracts caused a significant increase (P < 0.05) in the value of mean corpuscular hemoglobin concentration (MCHC) (Ogoloma et al., 2017). Adedosu et al. (2017) also reported a significant decrease in the counts of white blood cells (5.38 × 109/l), lymphocytes (2.24 × 109/l), and platelets (432 × 109/l) coupled with a significant (P < 0.05) increase in total serum proteins (9.5 mg/dl) and red blood cells (7.95 × 1012/l) in diabetic rats treated with ethanol leaf extract for 15 days. Another study conducted by Ogbonnaya et al. (2019) evaluated hematological indices of Wistar rats administered with aqueous and methanol leaf extracts of T. daniellii. There was a significant decrease in the hemoglobin concentration (13.70 g/l), red blood cells (62.20 × 106/μl), and packed cell volume (38.75%) in the group administered with 200 mg/kg methanol extract of T. daniellii as compared to the group administered with distilled water alone. However, there was no significant difference in the values obtained for rats administered with 200 mg/kg aqueous leaf extract of T. daniellii and distilled water alone. Thus, the authors concluded that the extracts had no significant effect on hematological indices.
Effect of T. daniellii on weight of treated rats
The effects of a 14-day oral dose of ethanol seed extracts (500, 1000, and 1500 mg/kg) on total body and organ weight were investigated and compared with the effects on ascorbic acid-treated rats (Shalom et al., 2017). There was no significant change in organ weights, but an observable reduction (16.33 g) in total body weight of the extract-treated animals was noted compared to the control animals (27.33 g). This might be due to the high crude fiber and low fat content of the seeds (Lim, 2012; Abiodun et al., 2014). Emudainohwo et al. (2015), however, reported a significant (7.76%) increase in the weight of diabetic rats treated with ethanol leaf extract of T. daniellii. Corroboratively, Nwonuma et al. (2016) reported a 1.70% weight increase in potassium bromate-exposed rats treated with ethanol leaf extract of T. daniellii.
Phytoremediative activity of T. daniellii
The use of plants in remediation and cleaning of petroleum-contaminated soils is cheap and very effective (Singh and Kumar, 2018). Plants such as Vicia faba and Zea mays have been documented to thrive and grow on soils polluted with hydrocarbons (Quinones-aquilar et al., 2003 ). Therefore, the ability of T. daniellii to grow on crude oil and acid-polluted soil was evaluated as a measure of its bioremediative capability and potential (Ekpo et al., 2012). In a 4-week study, different concentrations (0.5–17.5 ml) of crude oil were applied to the soil on which T. daniellii was grown. It was observed that morphological parameters such as mean chlorophyll content (2.51), leaf area (276.40), and number of leaves (4.7) of T. daniellii grown on crude oil were statistically nonsignificant from the control. Nonetheless, physiological parameters such as sulfate ion (0.3), aluminum oxide (0.3), hydrogen ion (0.4), and dry matter (91.6) contents were significantly different (Ekpo et al., 2012).
Insecticidal activity of T. daniellii
Ojekale et al. (2007) reported the insecticidal potential of the essential oil of T. daniellii leaves against Sitophilus zeamais. A lethal concentration of 345.2 mg per 100 ml after 72 h was reported. However, this observed activity was significantly inferior than those of standard insecticides (permethrin and allethrin).
Conclusions
Investigations of T. daniellii by various scientific studies revealed that the plant possesses far-reaching nutrients and medicinally important phytoconstituents. It can be deduced that the seeds have better nutritional values and importance. However, the methanol, acetone, and hexane extracts of the rhizome and leaves were observed to be toxic, and caution should be taken when consuming them. Conclusively, the aqueous and ethanol extracts of the seed and leaf were regarded as safe, while the ethanol leaf extract was found to be the most potent of the extracts of all morphological organs. Thus, it should be considered and used for its pharmaceutical potential.