

Introduction
Cancer is a persistent public health challenge facing society. As the second most common cause of death worldwide, cancer is expected to be the most significant obstacle to increasing life expectancy in the 21st century. The burden of cancer is increasing. In the coming decades, the greatest impact and fastest increase in the burden of cancer will continue to occur in developing countries, which not only causes immense suffering to patients but also imposes a heavy economic burden on society and patients’ families [1].
Since the occurrence and development of cancer is a complex and changeable process, various genetic changes may affect the treatment and prognosis of cancer patients [2, 3]. Pan-cancer analysis has been used by a wide range of researchers to explore the correlation between gene expression and clinical outcomes by analyzing large amounts of data from publicly funded databases. For example, previous studies have demonstrated that LAYN, SNLRP3, and SND1 could act as predictive biomarkers for cancer prognosis [4-6].
In recent years, immunotherapy has been a developing field in oncology [7]. The development of several novel immunomodulatory antibodies has altered the treatment of many cancers, to selectively enhance the host’s immune response to malignant diseases [8]. For instance, programmed death-1 (PD-1), an immune checkpoint molecule, has been recognized as a major target for cancer therapy [9]. Given the growing number of genetic abnormalities in tumors, immunotherapy appears to be a promising treatment option for cancer patients. In addition, tumor-infiltrating immune cells have been proven to influence the prognosis [10].
Protein tyrosine phosphatases (PTPs) are key regulators of cell activity, regulating tyrosine phosphorylation and signal transduction pathways [11]. The PTP family comprises 125 members, which can be divided into three categories: classical PTPs (acceptor and non-acceptor PTPs), bi-specific PTPs, and low molecular weight PTPs [12]. Changes in the delicate balance between tyrosine phosphorylation and dephosphorylation have been found to contribute to the pathogenesis of various genetic or acquired human diseases, including autoimmune, diabetes, and cancer [11, 13, 14].
Non-receptor protein tyrosine phosphatase 21 (PTPN21), also known as protein-tyrosine phosphatase D1 (PTPD1), is a cytoplasmic non-receptor tyrosine phosphatase in the PTP family. It was first isolated from human skeletal muscle by Møller et al. in 1994 [15]. Little research has been done on the function of PTPN21, but it has been shown to interact with epithelial and endothelial tyrosine kinases (Etk) of Tec kinase family members and to play a role in regulating cell growth and differentiation [16]. Meanwhile, some studies have found that PTPN21 may be involved in signal transduction of immune cell surface receptors, such as the T cell antigen receptor and Fas/CD95 [17-19]. In recent years, studies on the relationship between PTPN21 and human malignant tumors have also been increasing. Among them, PTPN21 can support the receptor stability and mitotic signal transduction of bladder cancer cells, and can also affect the development of gastric cancer by regulating the activity of STAT5 [20, 21]. Meanwhile, some researchers have found the presence of PTPN21 in the mutant gene of relapsed leukemia samples [22]. However, the research on PTPN21 in cancer is still in the early stage, and there are few studies on the treatment and prognosis of tumor patients. Therefore, this study aimed to look into the role of PTPN21 in pan-cancer and to summarize its effect on cancer prognosis and immune infiltration level.
Therefore, in this work, we aimed to explore the prognostic and immunological roles of PTPN21 in human cancers and confirm its oncogenic value in a variety of tumors.
Material and methods
PTPN21 expression in human cancers
First, we used the ONCOMINE (https://www.oncomine.org/) and TIMER2 databases (http://timer.cistrome.org/) to compare PTPN21 mRNA expression levels in different cancer types with their adjacent tissues [23, 24]. The conditions for setting the ONCOMINE database are as follows: p-value < 0.001, fold change > 2. The R language (version 3.6.3) and R package “ggplot2” were used to analyze and visualize the hard-processed RNAseq data of TCGA [25]. CPTAC analysis of the UALCAN database (http://ualcan.path.uab.edu/analysis-prot.html) was used to detect the protein expression level of PTPN21 in glioblastoma multiforme, renal cell carcinoma, lung adenocarcinoma, and hepatocellular carcinoma [26]. The Human Protein Atlas (HPA) (http://www.proteinatlas.org/) was used to demonstrate the expression of PTPN21 in normal and tumor tissues of the kidney and lung [27].
Prognostic survival analysis
The TCGA and GEO datasets were employed to investigate the correlation of PTPN21 expression with the prognosis of different cancers. GEPIA2 (http://gepia2.cancer-pku.cn/) was adopted to analyze the prognostic correlation between TCGA expression of PTPN21 in a variety of tumors [28]. The Kaplan-Meier Plotter database (https://kmplot.com/analysis/) was used to analyze the correlation between PTPN21 expression and overall survival (OS) [29].
Genetic alteration analysis
The cBioPortal database (https://www.cbioportal.org/) was used to analyze changes in the PTPN21 gene [30]. First, we calculated the mutation frequency and copy number changes of the PTPN21 gene based on the TCGA universal cancer atlas dataset and created the mutation sites map of PTPN21. In addition, the correlation between PTPN21 mutation status and prognosis of uterine corpus endometrial carcinoma (UCEC) was analyzed according to “uterine endometrial carcinoma (TCGA generalized carcinoma)”.
Immune infiltration analysis
In this study, we analyzed the correlation between PTPN21 expression in bladder urothelial carcinoma (BLCA), kidney renal clear cell carcinoma (KIRC), and lung squamous cell carcinoma (LUSC) and tumor immune infiltration cells in TIMER2, including B cells, CD8+ T cells, CD4+ T cells, macrophages, neutrophils, and dendritic cells (DCs). To further evaluate the role of PTPN21 expression in tumor immune infiltration, we used TIMER2 to examine the relationship between PTPN21 and various prognostic immune cell markers to identify potential infiltrating immune cell subtypes. These immune cells include B cells, T cells (general), CD8+ T cells, helper T cells (Tfh), T-helper 1 (Th1) cells, T-helper 2 (Th2) cells, T-helper 9 (Th9) cells, T-helper 17 (Th17) cells, T-helper 22 (Th22) cells, Tregs, exhausted T cells, M1 macrophages, M2 macrophages, tumor-associated macrophages (TAMs), monocytes, natural killer (NK) cells, neutrophils, and DCs.
Enrichment analysis of PTPN21-related genes
The R packages “ggplot2” and “clusterProfiler” were used to mine the co-expressed genes of PTPN21. Meanwhile, GO and KEGG enrichment analyses were analyzed and visualized.
Statistical analysis
The differential expression of PTPN21 in cancer tissues and normal tissues was analyzed in the ONCOMINE database using the p-value, fold change, and gene grade. The survival curves were drawn by Kaplan-Meier Plotter and GEPIA2 databases. Spearman correlation was used to analyze the correlation between PTPN21 and immune cells and their markers in TIMER2. In this study, p < 0.05 was considered statistically significant.
Results
PTPN21 expression in pan-cancer
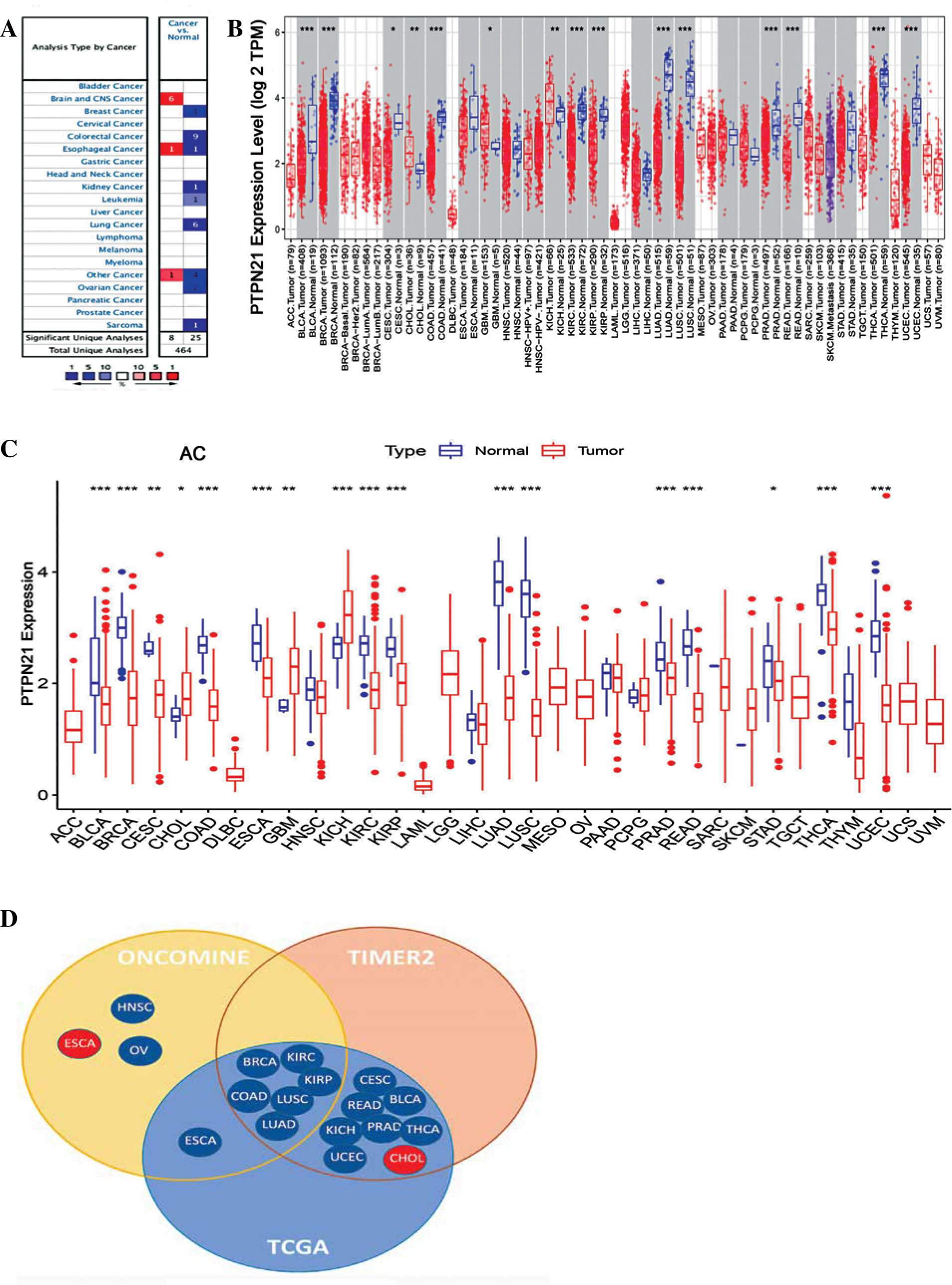
We analyzed PTPN21 mRNA expression levels in tumors and adjacent tissues using ONCOMINE, TIMER2, and TCGA databases. In ONCOMINE, PTPN21 expression was higher in the brain and central nervous system (CNS) cancer than in normal tissues. Conversely, PTPN21 mRNA expression was reduced in breast, colorectal, kidney, leukemia, sarcoma, lung, and ovarian cancer. Notably, PTPN21 expression was higher in one dataset and lower in the other in esophageal cancer compared with normal tissue (Fig. 1A). By searching in the TIMER2, the expression level of PTPN21 was found significantly down-regulated in many types of cancers, including bladder urothelial carcinoma (BLCA), breast invasive carcinoma (BRCA), colon adenocarcinoma (COAD), kidney renal clear cell carcinoma (KIRC), kidney renal papillary cell carcinoma (KIRP), lung adenocarcinoma (LUAD), lung squamous cell carcinoma (LUSC), prostate adenocarcinoma (PRAD), rectum adenocarcinoma (READ), thyroid carcinoma (THCA), and uterine corpus endometrial carcinoma (UCEC). However, it was significantly increased in cholangiocarcinoma (CHOL), glioblastoma multiforme (GBM), and kidney chromophobe (KICH) (Fig. 1B). The tumor and normal tissues for PTPN21 across all TCGA tumors are shown in Figure 1C. Figure 1D shows a comparative analysis of PTPN21 expression in tumors from ONCOMIE, TIMER2, and TCGA databases.
Fig. 1
Expression levels of PTPN21 in different cancer types. A) The ONCOMINE database showed that PTPN21 expression increased or decreased compared to normal tissues of different cancer types (the numbers in the grid represent the number of included cases; red means higher, blue means lower; the darker the color, the greater the difference in expression). B) Expression levels of PTPN21 in different tumor types were analyzed using the TIMER2 database. C) Expression of PTPN21 in pan-cancer was analyzed using paired tumor/ normal samples from TCGA database. D) Comparison summary of the above three databases. *p < 0.05, **p < 0.01, ***p < 0.001
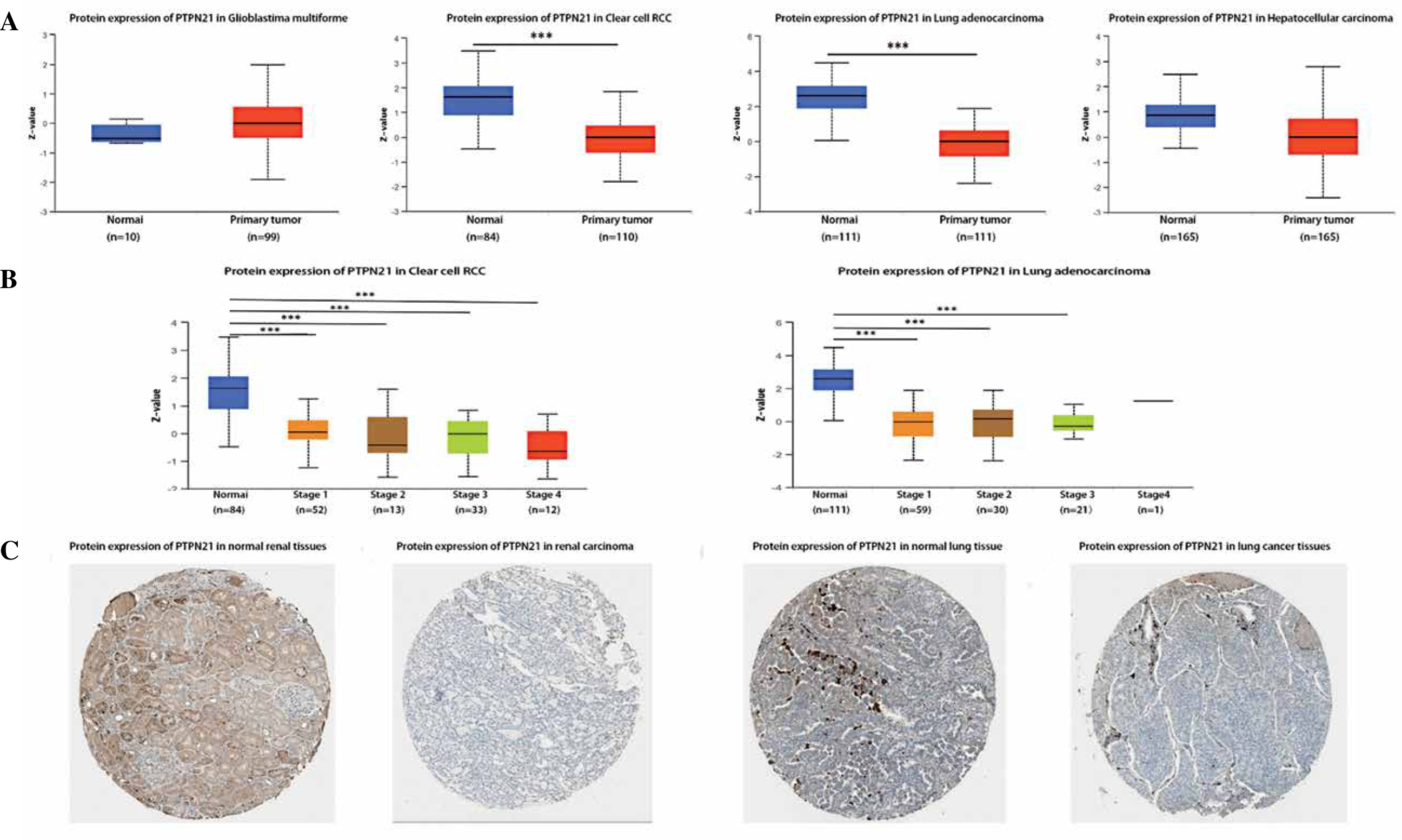
We further confirmed that PTPN21 expression was significantly reduced in BLCA, BRCA, CESC, COAD, ESCA, KIRC, KIRP, LUAD, LUSC, PRAD, READ, THCA, and UCEC. Moreover, CPTAC analysis showed that PTPN21 protein expression was down-regulated and correlated with pathological stages of clear cell RCC and LUAD (Fig. 2A, B). The data of the HPA database showed the different expression levels of PTPN21 in normal or tumor tissue of the kidney and lung (Fig. 2C). In a word, PTPN21 was low expressed in most cancers.
Fig. 2
Expression of PTPN21 protein in different cancers. A) CPTAC was used to analyze PTPN21 protein expression in glioblastoma multiforme, renal cell RCC, lung adenocarcinoma, and hepatocellular carcinoma. B) Expression of PTPN21 protein was analyzed by the main pathological stages of clear cell RCC and lung adenocarcinoma. C) Based on the HPA database, the expression of PTPN21 protein in normal and tumor tissues of kidney and lung was demonstrated. *p < 0.05, **p < 0.01, ***p < 0.001
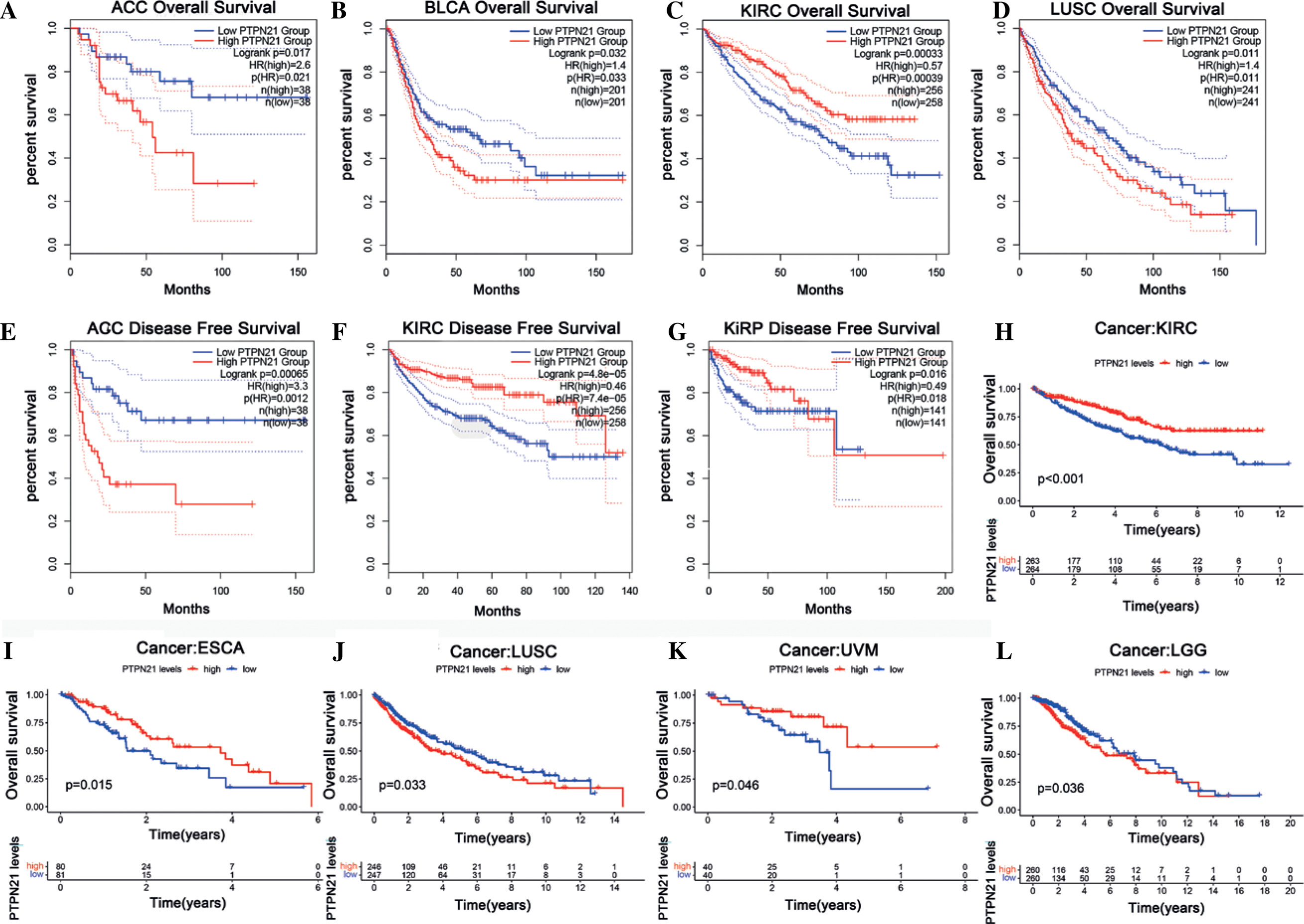
Prognostic value of PTPN21 in cancers
First, GEPIA2 was used to analyze the prognostic value of PTPN21 in different tumor types in the TCGA data set. High PTPN21 expression was associated with poorer OS in adrenocortical carcinoma (ACC) (p = 0.017), BLCA (p = 0.032), and LUSC (p = 0.011), but with a better OS in KIRC (p = 0.00033) (Fig. 3A-D). In addition, DFS analysis showed that high PTPN21 expression was a marker of poor prognosis in patients with ACC (p = 0.00065), but a marker of good prognosis in patients with KIRC (p = 0.00065) and KIRP (p = 0.00065) (Fig. 3E-G). Based on the TCGA databases, survival analysis results demonstrated that PTPN21 expression significantly affects prognosis in KIRC, ESCA, LUSC, uveal melanoma (UVM), and lower grade glioma (LGG) (Fig. 3H-L). These results suggested that increased PTPN21 expression was associated with better patient outcomes across multiple tumor types.
Fig. 3
Correlation between PTPN21 expression and prognostic survival in patients with various TCGA tumor types. A-D) Kaplan-Meier plot of significant overall survival (OS) results in GEPIA2. E-G) Kaplan-Meier plot of significant disease-free survival (DFS) results in GEPIA2. H-L) Overall prognosis survival curve of the TCGA database
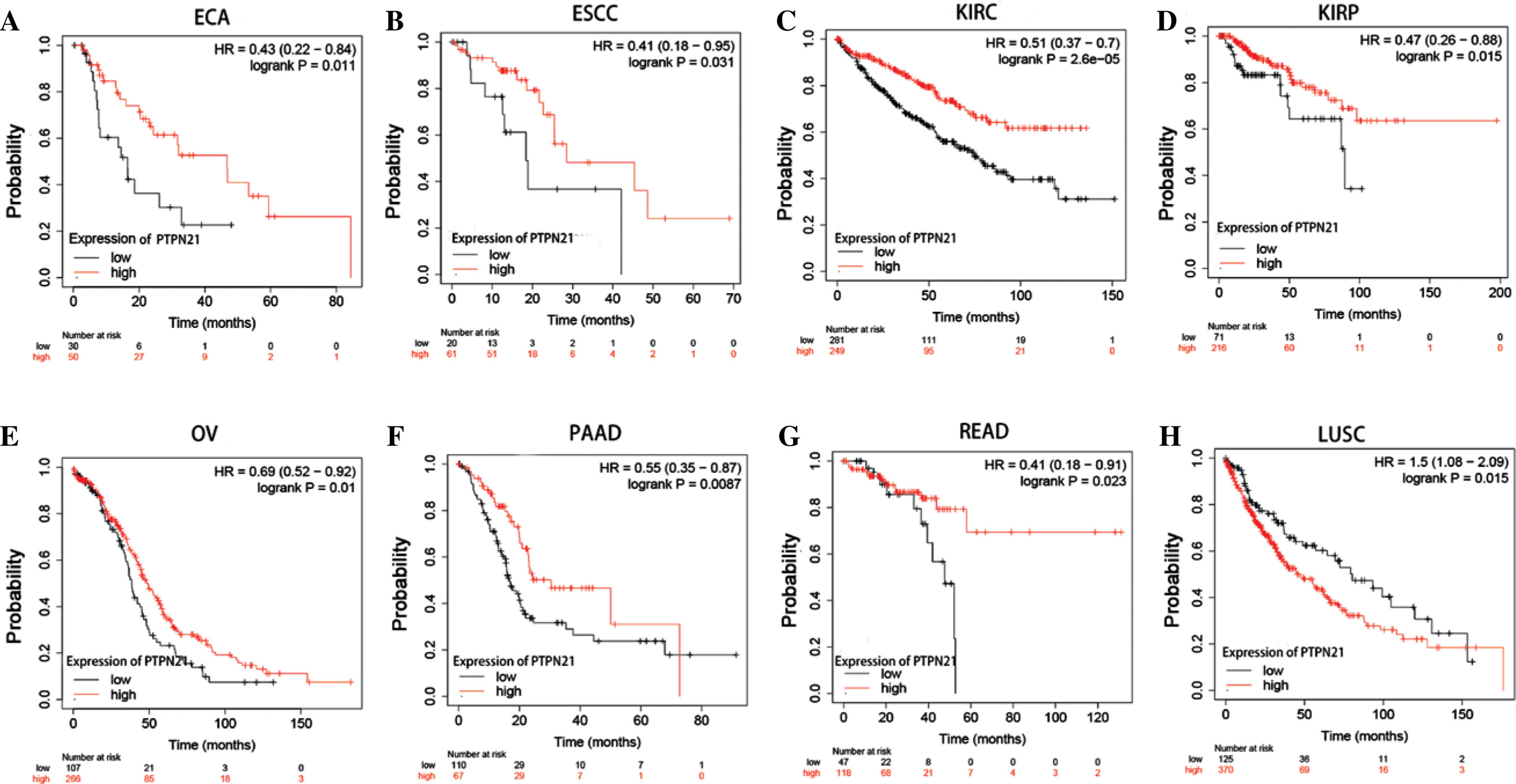
In addition, we used the Kaplan-Meier Plotter database containing TCGA, GEO and ECA datasets to analyze the correlation between PTPN21 expression and prognosis in different tumor patients. In esophageal adenocarcinoma (ECA) (p = 0.011), esophageal squamous cell carcinoma (ESCA) (p = 0.031), KIRC (p = 2.6E-05), KIRP (p = 0.015), ovarian cancer (OV) (p = 0.01), pancreatic adenocarcinoma (PAAD) (p = 0.0087), and READ (p = 0.023), upregulation of PTPN21 was associated with better OS in patients, but it was associated with worse OS in LUSC (p = 0.015) (Fig. 4A-H).
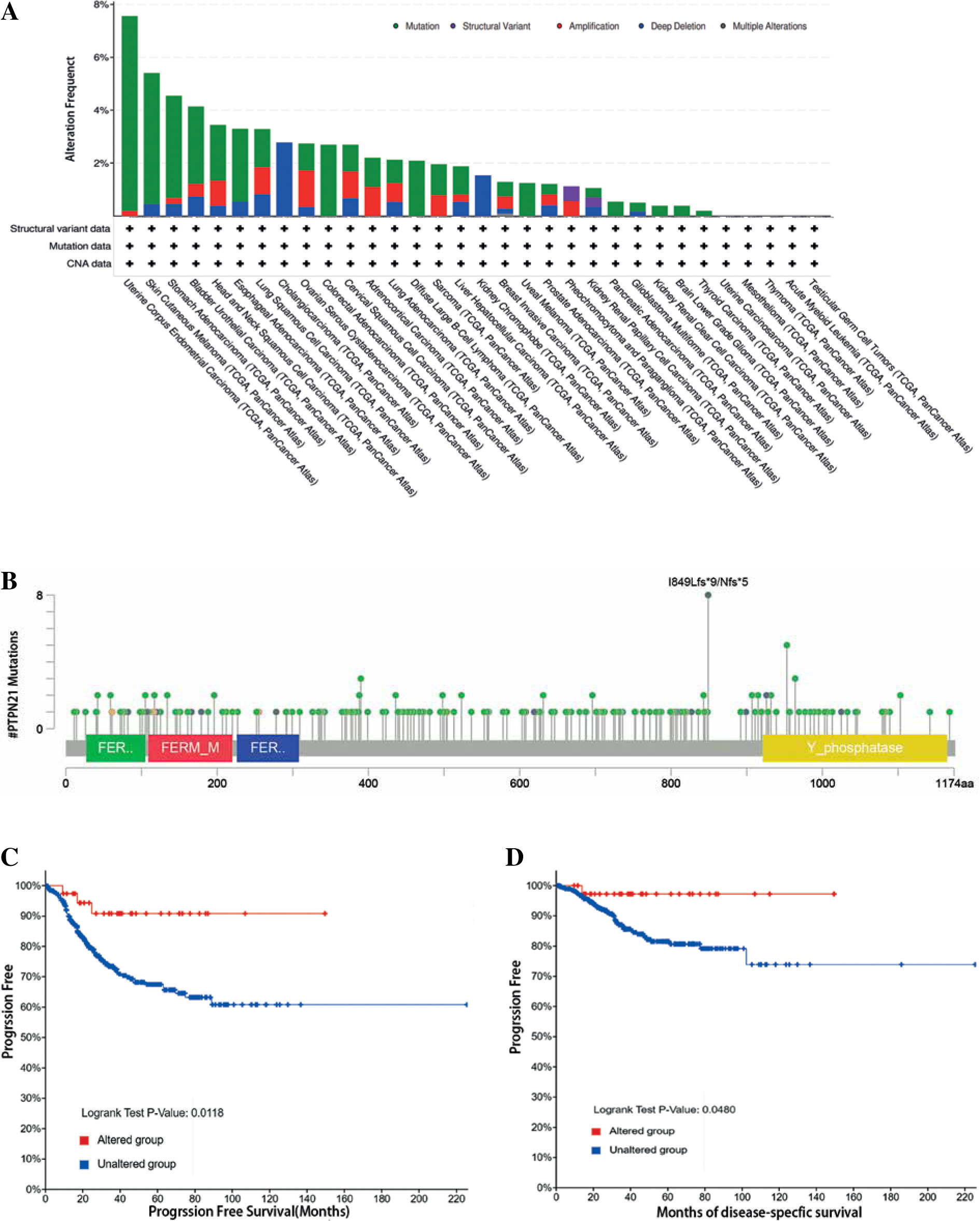
Genetic mutation analysis
We studied the genetic mutation of PTPN21 in different tumor types in the TCGA dataset using the cBioPortal database. The UCEC tumor samples were found to have the highest PTPN21 genetic alteration frequency (> 6%) (Fig. 5A). Mutations and copy number amplification were the most common genetic changes found in UCEC tumor samples, with mutations predominating. Mutations were also the dominant type of genetic change in all TCGA tumor samples. As shown in Figure 5B, PTPN21 mutations were detected in TCGA tumor samples, including missense mutation, truncating mutation, splice mutation, fusion mutation, and inframe mutation. Following that, we explored the link between PTPN21 gene alteration and clinical outcomes in cancer patients. As shown in Figures 5C and D, PTPN21 mutations were associated with a favorable prognosis of progression-free survival (PFS) (p = 0.0118) and DFS (p = 0.0480) in UCEC patients.
Fig. 5
PTPN21 genetic alteration in various tumor types of TCGA. A) Types of PTPN21 gene alterations produced by the cBioPortal database. B) Mutation site of PTPN21. C, D) cBioPortal analyzed the association between PTPN21 mutation status and progression-free survival and disease-specific survival in UCEC
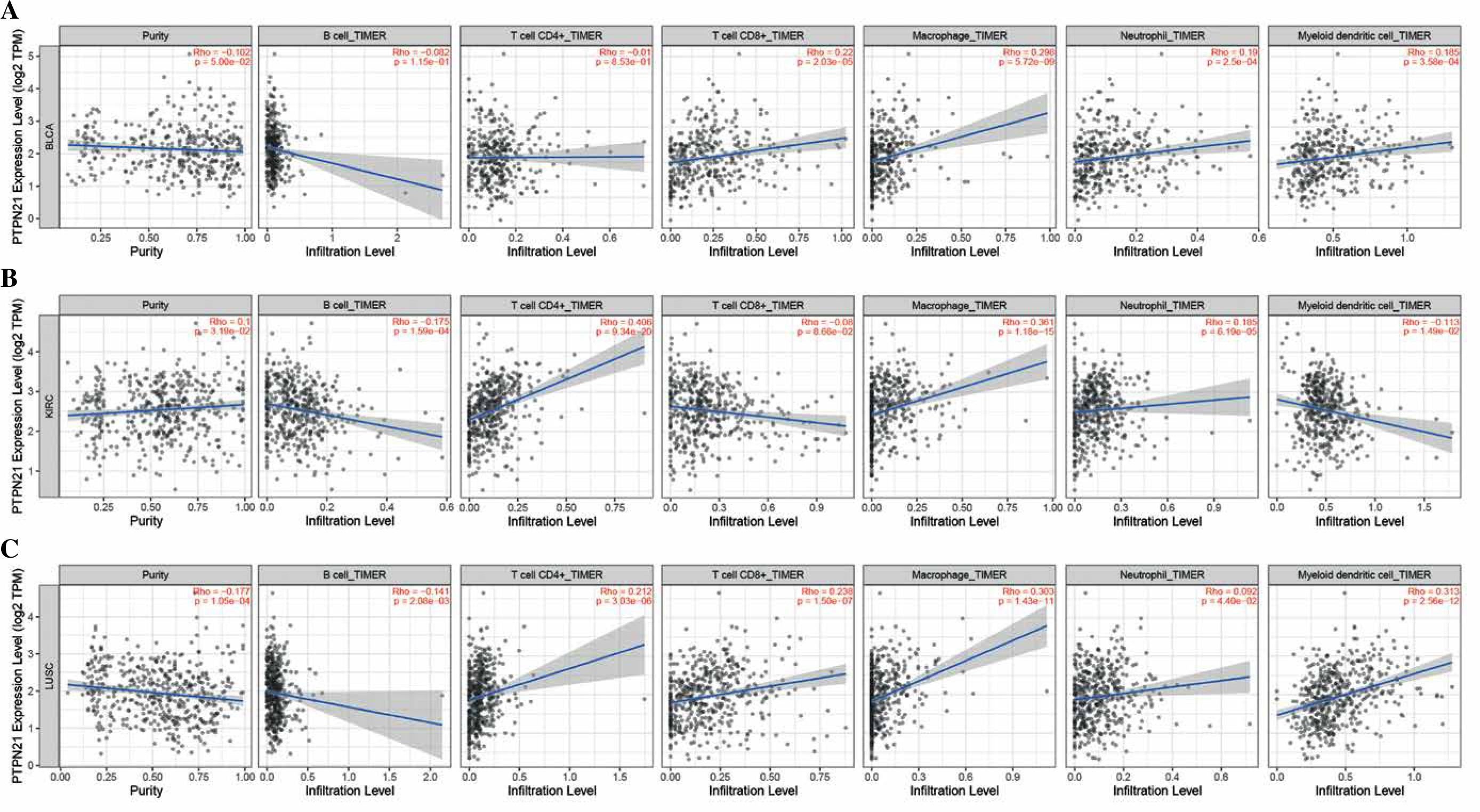
Immune cell infiltration analysis
More and more research has shown that immune cell infiltration in the tumor microenvironment can affect cancer cell survival, local invasion, and metastasis [31, 32]. Therefore, we used the TIMER2 database to explore the correlation between PTPN21 expression and the level of infiltration of six immune cells in cancer. Considering that the expression of PTPN21 in a variety of cancers is related to the level of immune infiltration, we conducted a specific analysis combined with immune infiltration and prognosis. We selected cancer types whose PTPN21 expression was negatively correlated with tumor purity in TIMER2 and strongly associated with poor prognosis in GEPIA2, namely BLCA, KIRC, and LUSC. As shown in Figure 6A, PTPN21 expression was associated with CD8+ T cells (R = 0.22, p = 2.03e-05), macrophages (R = 0.298, p = 5.72e-09), neutrophils (R = 0.19, p = 2.5e-04) and DCs (R = 0.185, p = 3.58e-04) in BLCA. The expression of PTPN21 in KIRC was significantly positively correlated with B cells (R = –0.175, p = 1.59e-04), CD4+ T cells (R = 0.406, p = 9.34e-20), macrophages (R = 0361, p = 1.18e-15), neutrophils (R = 0.185, p = 6.19e-05), and DCs (R = –0.113, p = 1.49e-02) (Fig. 6B). In LUSC, PTPN21 expression was positively correlated with infiltration of B cells (R = –0.141, p = 2.08e-03), CD4+ T cells (R = 0.212, p = 3.03e-06), CD8+ T cells (R = 0.238, p = 1.50e-07), macrophages (R = 0.303, p = 1.43e-11), neutrophils (R = 0.092, p = 4.40e-02), and DCs (R = 0.313, p = 2.56e-12) (Fig. 6C). However, PTPN21 expression was not associated with B cell infiltration in BLCA and CD8+ T cell infiltration in KIRC. In conclusion, PTPN21 expression was significantly correlated with immune infiltration in tumors, suggesting that PTPN21 may influence cancer patient survival by influencing immune infiltration in the tumor microenvironment.
Fig. 6
Correlation of PTPN21 expression with immune invasion levels in BLCA, KIRC, and LUSC. A) PTPN21 expression was significantly positively correlated with the infiltration levels of CD8+ T cells, macrophages, neutrophils and dendritic cells (DCs) in BLCA. B) PTPN21 expression was significantly positively correlated with the infiltration levels of B cells, CD4+ T cells, macrophages, neutrophils and DCs in KIRC. C) Expression of PTPN21 was positively correlated with infiltration levels of B cells, CD4+ T cells, CD8+ T cells, macrophages, neutrophils and DCs in LUSC
Immune markers correlation analysis
In addition to the six immune infiltrating cells described above, we used the TIMER2 database to examine the relationship between PTPN21 expression in BLCA, KIRC, LUSC, and other immune cell markers. As shown in Table 1, after purity adjustment, PTPN21 expression was closely associated with 28 immune cell markers in BLCA, 42 in KIRC, and 36 in LUSC.
Table 1
Correlation analysis between PTPN21 and markers of immune cells in TIMER2
In addition to the above overall changes, PTPN21 was significantly associated with immune markers of several T cells in BLCA, including Th2 (STAT6 and CCR4), Th9 (TGFBR2), Th17 (IL1R1 and STAT3), Th22 (AHR), and Treg (CCR8 and FOXP3). Meanwhile, PTPN21 was also closely associated with the levels of most expression markers of TAMs, M1 macrophages, M2 macrophages, monocytes, neutrophils, and DCs, including CL2, IL10, COX2, CD163, MS4A4A, CD115, CD11b, CD15, BDCA-4, and CD11c. However, there was no correlation between PTPN21 and immune markers of B cells, CD8+ T cells, and NK cells. Compared with BLCA, markers of B cells, T regulatory cells, CD8+ T cells, exhausted T cells, and NK cells were significantly correlated with the expression of PTPN21 in KIRC. In addition, in LUSC, PTPN21 was strongly correlated with T cells with different functions, such as Th1, Th2, Th9, Th17, Th22, and Treg. At the same time, it was correlated with M1 macrophages, M2 macrophages, neutrophils, and DCs. In conclusion, the expression of PTPN21 in BLCA, KIRC, and LUSC was associated with different degrees of immune cell infiltration in different ways. These analyses further confirmed that PTPN21 expression plays an important role in the immune infiltration of cancer.
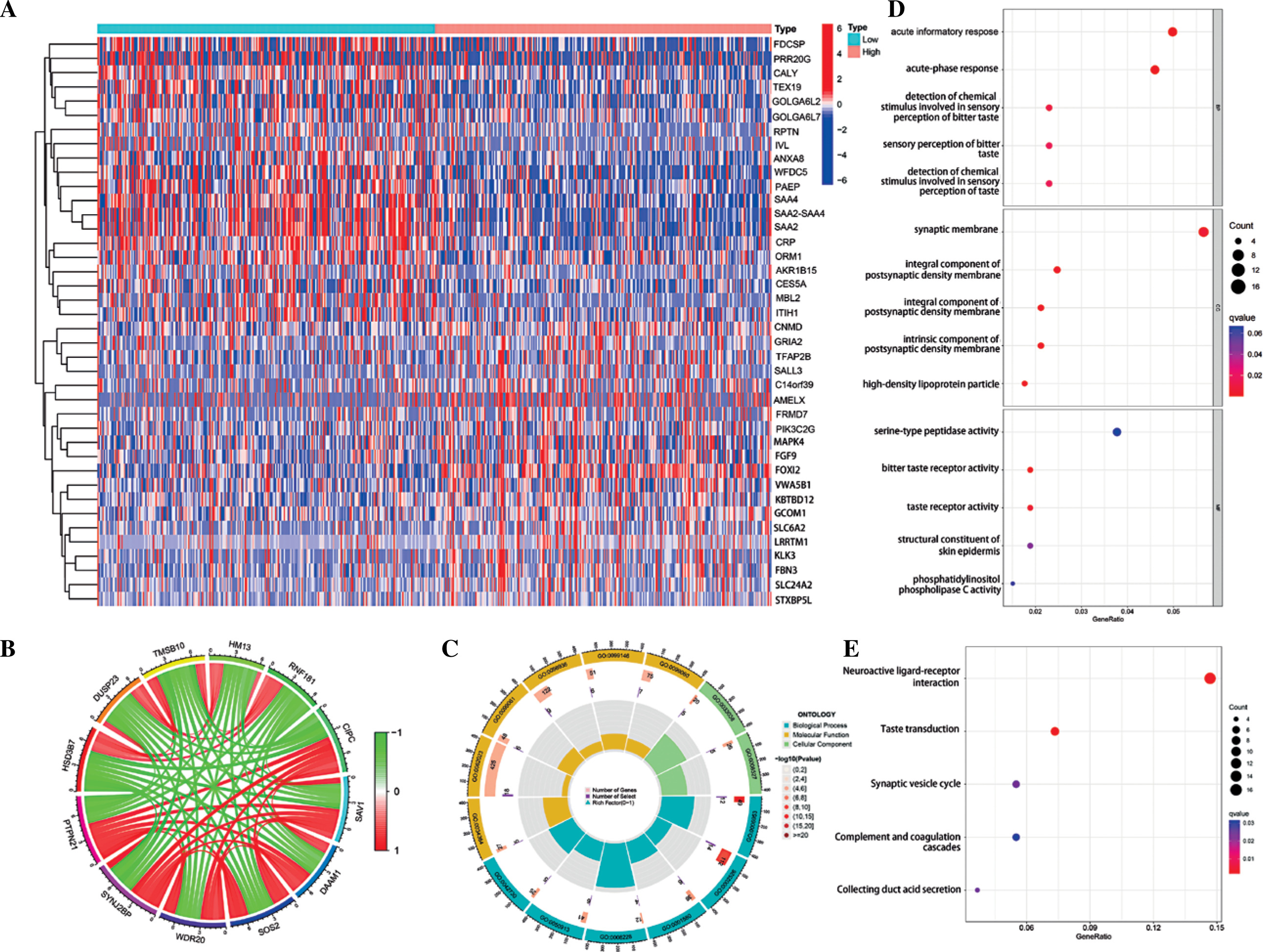
Enrichment analysis of PTPN21-related genes
Heat maps and circles show PTPN21 and its major related genes (Fig. 7A, B). GO analysis showed that PTPN21 and PTPN21-binding proteins were mainly involved in biological processes including regulation of the acute informatory response and acute-phase response, detection of a chemical stimulus involved in sensory perception of bitter taste, and sensory perception of bitter taste. Meanwhile, the cellular components involved in PTPN21 include synaptic membrane, integral component of postsynaptic specialization membrane, and integral component of postsynaptic density membrane. The molecular functions involved include serine-type peptidase activity, bitter taste receptor activity, taste receptor activity, and structural constituent of skin epidermis. (Fig. 7C, D). KEGG analysis showed that PTPN21 and its related proteins were mainly concentrated in neuroactive ligand-receptor interaction, taste transduction, and synaptic vesicle cycle pathways (Fig. 7E).
Discussion
In this research, we comprehensively investigated the molecular characteristics of PTPN21 in 33 different cancers from different databases, including TCGA, GEO, and CPTAC, aiming to investigate the value of PTPN21 in cancer prognosis, progression, and treatment.
In our study, PTPN21 expression was down-regulated in a variety of tumor tissues, especially in BLCA, KIRC, KIRP, LUAD, LUSC, and READ. In addition, the UALCAN database confirmed that the protein expression level of PTPN21 in KIRC and LUAD tumor tissues was low and normal, and the high expression of PTPN21 was associated with the early clinical stage.
We further explored the relationship between PTPN21 expression and clinical parameters or prognosis. The GEPIA2 database survival analysis revealed that high PTPN21 expression was associated with better prognostic survival in KIRC and KIRP, but not in ACC, BLCA, or LUSC. Survival analysis of the TCGA database showed that PTPN21 expression significantly affected the prognosis of KIRC, ESCA, LUSC, UVM, and LGG. Additional data sources are available in the Kaplan-Meier Plotter database, and prognostic survival analysis revealed that high PTPN21 expression was associated with longer prognostic survival in patients with OV, COAD, ECA, ESCC, KIRC, KIRP, and PAAD.
There is increasing evidence that genomic mutations are associated with tumor progression and chemotherapy response. For example, Hollis et al. found that BRCA1 and BRCA2 mutations were significantly associated with patient survival, which may be the result of a significant response to platinum therapy [33]. In addition, it has been found that mutations of TP53 often occur at specific metastatic sites, which may be biomarkers or therapeutic targets for patients with metastatic breast cancer [34]. This study found that PTPN21 mutations were most common in UCEC. We analyzed whether PTPN21 mutations affect clinical outcomes in cancer patients, and found that PTPN21 mutations may have a protective effect on UCEC patients. Taken together, these findings suggest that PTPN21 plays a role in the progression of multiple cancers and is a promising predictor of practical application in cancer prognosis.
It is important to note that another key part of this study is that PTPN21 expression is associated with levels of various immune infiltrations in cancer. We found significant associations between PTPN21 and infiltration of CD4 T+ cells, macrophages, neutrophils, and DCs in BLCA, KIRC, and LUSC. With the development of immune checkpoint inhibitors in recent years, immune cell biomarkers can now be used as prognostic markers as well as a new therapeutic approach [35]. Therefore, we used TIMER2 to evaluate the association between multiple immune cell markers in BLCA, KIRC, and LUSC and PTPN21 expression. PTPN21 was closely associated with CL2, IL10, COX2, CD163, MS4A4A, CD115, CD11b, CD15, BDCA-4, and CD11c in BLCA markers of TAMs, M1 macrophages, M2 macrophages, neutrophils, and DCs. In addition, PTPN21 was found to regulate macrophage polarization and DC infiltration in LUSC and KIRC. These findings suggest that PTPN21 may play a regulatory role in TAM polarization, which has been revealed to promote tumor growth by inhibiting immune clearance, promoting tumor cell proliferation, and stimulating angiogenesis [29]. Meanwhile, PTPN21 was strongly correlated with B cells, Th1 cells, Th2 cells, Th9 cells, Th17 cells, Th22 cells, and Treg. In conclusion, PTPN21 can affect the invasion level of most immune cells in cancer, but how it affects the occurrence and development of cancer needs further study.
Finally, we identified several genes co-expressed with PTPN21 in tumor tissues. GO analysis showed that these genes were mainly located in the synaptic membrane, integral component of postsynaptic specialization membrane, and integral component of postsynaptic density membrane. At the same time, it is involved in the acute informatory response, the acute-phase response, detection of a chemical stimulus involved in sensory perception of bitter taste and sensory perception of bitter taste, and molecular functions such as serine-type peptidase activity, bitter taste receptor activity, taste receptor activity and a structural constituent of skin epidermis. In addition, KEGG enrichment analysis showed that PTPN21 and its related genes mainly regulate neuroactive ligand-receptor interaction, taste transduction, and the synaptic vesicle cycle. The mechanism mentioned above has not been studied in cancer, so further studies are needed to verify it.
The study had some limitations. First, some of the less common tumor types had small sample sizes, which could result in batch effects or inaccurate results. Secondly, only online databases were used in this study to preliminarily link PTPN21 with the progression of a variety of tumors, and more in vivo or in vitro experiments are needed to verify the precise molecular functions of PTPN21 in tumorigenesis.
In conclusion, PTPN21 expression is decreased in a variety of cancer tissues, and its expression and gene changes are statistically correlated with the clinical outcomes of some tumor patients. In addition, immune infiltration analysis provides a potential mechanism by which PTPN21 regulates tumor immunity. Therefore, further experimental and clinical studies are needed to investigate the practical application of PTPN21 in cancer treatment and prognostic prediction.